










-
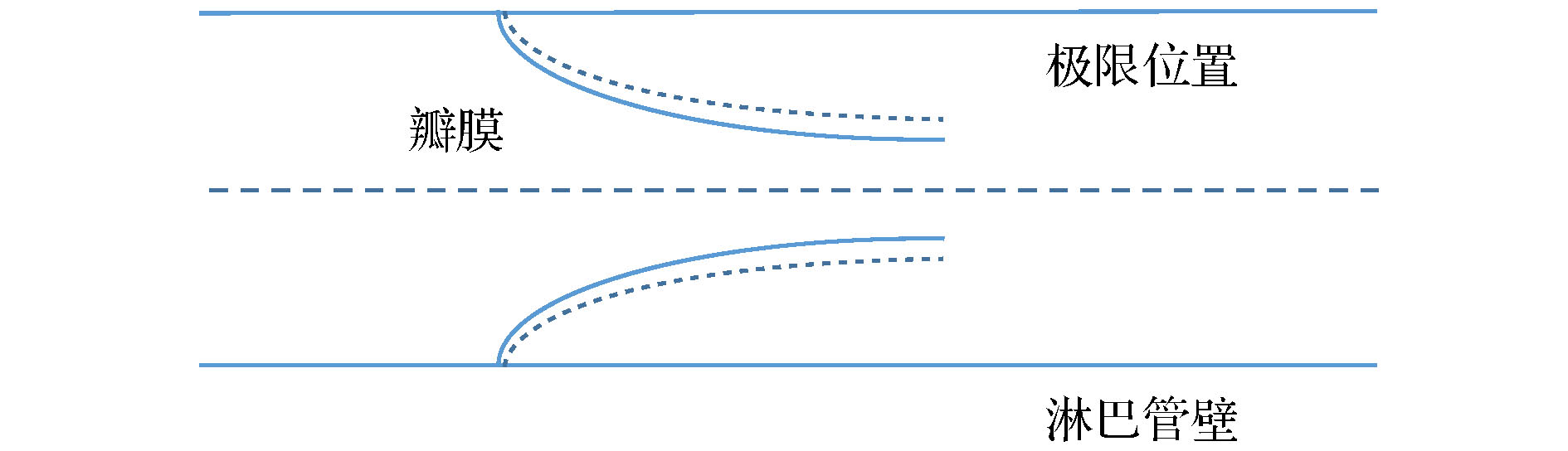
淋巴系统是人体内重要的防御功能系统, 具有三大免疫功能, 首先是能够抵御细菌病毒, 使人体免于疾病的攻击; 其次是由淋巴细胞加以辅助, 清除由新陈代谢而出的产物; 最后是由淋巴细胞来修补受损的器官与组织, 使其恢复正常的生理功能。淋巴系统没有像血液循环系统中心脏一样的动力泵, 淋巴液的驱动主要靠淋巴管的自主收缩来完成(肺淋巴系统是靠肺泡的运动)。淋巴管的自主收缩循环是由淋巴肌细胞内钙离子增加产生收缩, 收缩驱动流体产生剪切力, 剪切力使淋巴内皮细胞产生一氧化氮合酶(eNOS), 一氧化氮合酶使一氧化氮增加, 一氧化氮的增加降低钙离子使淋巴管松弛, 淋巴管松弛后流体剪切率下降, eNOS下降, 一氧化氮下降, 钙离子增加, 淋巴肌细胞收缩, 开始新的周期。可见一氧化氮的浓度及其分布对淋巴管的收缩起关键作用。显然出口压力会影响淋巴管内流体的剪切率, 进而影响一氧化氮的浓度和淋巴管的收缩。为了研究淋巴管出口压力对淋巴管收缩的影响, 建立了一个晶格玻尔兹曼模型, 模拟嵌入多孔组织的初始淋巴管和有两对瓣膜的集合淋巴管, 该模型可以重现一氧化氮、钙的相互影响以及淋巴管的自主收缩, 并研究不同出口压力下一氧化氮的分布及其平均值.The lymphatic system is an important defense function system in the human body. It is also critical to humoral homeostasis. Local dysfunction will cause edema, immune deficiency, and a high incidence. There are intraluminal valves in the lymphatic system, which allows the lymph fluid to flow to the large veins and heart. It has three major immune functions. First, it can resist bacterial viruses and protect the human body from disease attacks. Secondly, it is supplemented by lymphocytes to remove the products produced by metabolism. In the end, The damaged organs and tissues are repaired by lymphocytes to restore normal physiological functions. The lymphatic system does not have the same pump as the heart of the blood circulatory system. The driving of lymph is mainly done by the spontaneous contraction of the lymphatics (the lung lymphatic system is compressed by the alveoli). The autonomic contraction cycle of lymphatic vessels is caused by the increase of Ca2+ in lymphocytes, and the contraction drives the fluid to produce shearing force. The shearing force produces nitric oxide synthase (eNOS) in lymphatic endothelial cells, and eNOS increases NO and increases NO. Decreasing Ca2+ relaxes lymphatic vessels, fluid shear rate decreases after lymphatic vessel relaxation, eNOS decreases, NO decreases, Ca2+ increases, lymphocytes contract, and a new cycle begins. It can be seen that the concentration of NO and its distribution play a key role in the contraction of lymphatic vessels. Obviously, export pressure affects the shear rate of fluid in the lymphatics, which in turn affects the concentration of NO and the contraction of lymphatic vessels. To investigate the effect of lymphatic outlet pressure on lymphatic vessel contraction, we established a lattice Boltzmann model to simulate the initial lymphatic vessels embedded in porous tissue and the collecting lymphatic vessels with two pairs of valves. The valve is the main source of NO. Once contraction begins, the contraction is spontaneous, self-sustaining, and the system exhibits non-linear dynamics. This model can reproduce NO and The interaction of Ca2+ and the spontaneous contraction of lymphatic vessels, and the distribution of NO under different outlet pressures and their average values were studied.
-
Keywords:
- Lymphatic vessel /
- lattice Boltzmann method /
- valve /
- NO
[1] Louveau A, Smirnov I, Keyes T J, Eccles J D, Rouhani S J, Peske J D, Derecki N C, Castle D, Mandell J W, Lee K S, Harris T H, Kipnis J 2015 Nature 523 377
[2] Margaris K N, Black R A 2012 J. R. Soc. Interface 9 601
 Google Scholar
Google Scholar
[3] Macdonald A J, Arkill K P, Tabor G R, McHale N G, Winlove C P 2008 Am. J. Physiol. Heart C. 295 305
Google Scholar
[4] 张立民 2012 ATP敏感性钾通道在一氧化氮调节失血性休克大鼠离体淋巴管泵功能中的作用 (张家口: 河北北方学院)
Zhang L M 2012 Role of ATP-Sensitive Potassium Channels in Nitric Oxide in Regulating the Function of Isolated Lymphatic Pump in Hemorrhagic Shock(Zhangjiakou: Hebei North University) (in Chinese)
[5] 秦立鹏, 牛春雨, 赵自刚 2011 生理科学进展 42 237
Qin L P, Niu C Y, Zhao Z G 2011 Advances in Physiological Sciences 42 237
[6] Kunert C, Baish J W, Liao S, Padera T P, Munn L L 2015 PNAS 112 10938
Google Scholar
[7] Baish J W, Kunert C, Padera T P, Munn LL 2016 PLoS Comput. Biol. 12 1005
[8] 赵彤彤 2018 多孔介质含天然气水合物多相流动LBM模拟 (太原: 太原理工大学)
Zhao T T 2018 LBM Simulation of Multiphase Flow of Natural Gas Hydrate in Porous Media (Taiyuan: Taiyuan University of Technology) (in Chinese)
[9] Shan X, Chen H 1993 Phys. Rev. E 47 1815
Google Scholar
[10] Li H B, Mei Y M, Maimon N, Padera T P, Baish J W, Munn L L 2019 SCIENTIFICREPORTS 9 2045
[11] Chen, Chen, Martnez, Matthaeus 1991 Phys. Rev. Lett. 67 27
[12] Qian Y H, D’HumièresD, Lallemand P 1992 Europhys. Lett. 17 479
Google Scholar
[13] Sukop M C, ThorneJr D T2010 Lattice Boltzmann Modeling: An Introduction for Geoscientists and Engineers (Berlin: Springer Publishing Company) p36
[14] Pujol F, Hodgson T, Martinezcorral I, Prats A C, Devenport D, Takeichi M, Genot E, Mäkinen T, Francis-West P, Garmy-Susini B, Tatin F 2017 Arterioscl. Thromb. Vas. Biol. 37 1732
Google Scholar
[15] Scallan J P, Davis M J 2013 J. Physiol. 591 250
[16] Kawai Y, Yokoyama Y, Kaidoh M 2010 Am. J. Physiol. 298 647
Google Scholar
[17] Ladd A J C, Verberg R 2001 J. Stat. Phys. 104 1191
Google Scholar
[18] He X, Doolen G 1997 J. Comput. Phys. 134 306
Google Scholar
[19] H Glenn B, Olga Yu G, Zawieja D C 2011 Ame. J. Physiol. Heart C. 301 1897
Google Scholar
-
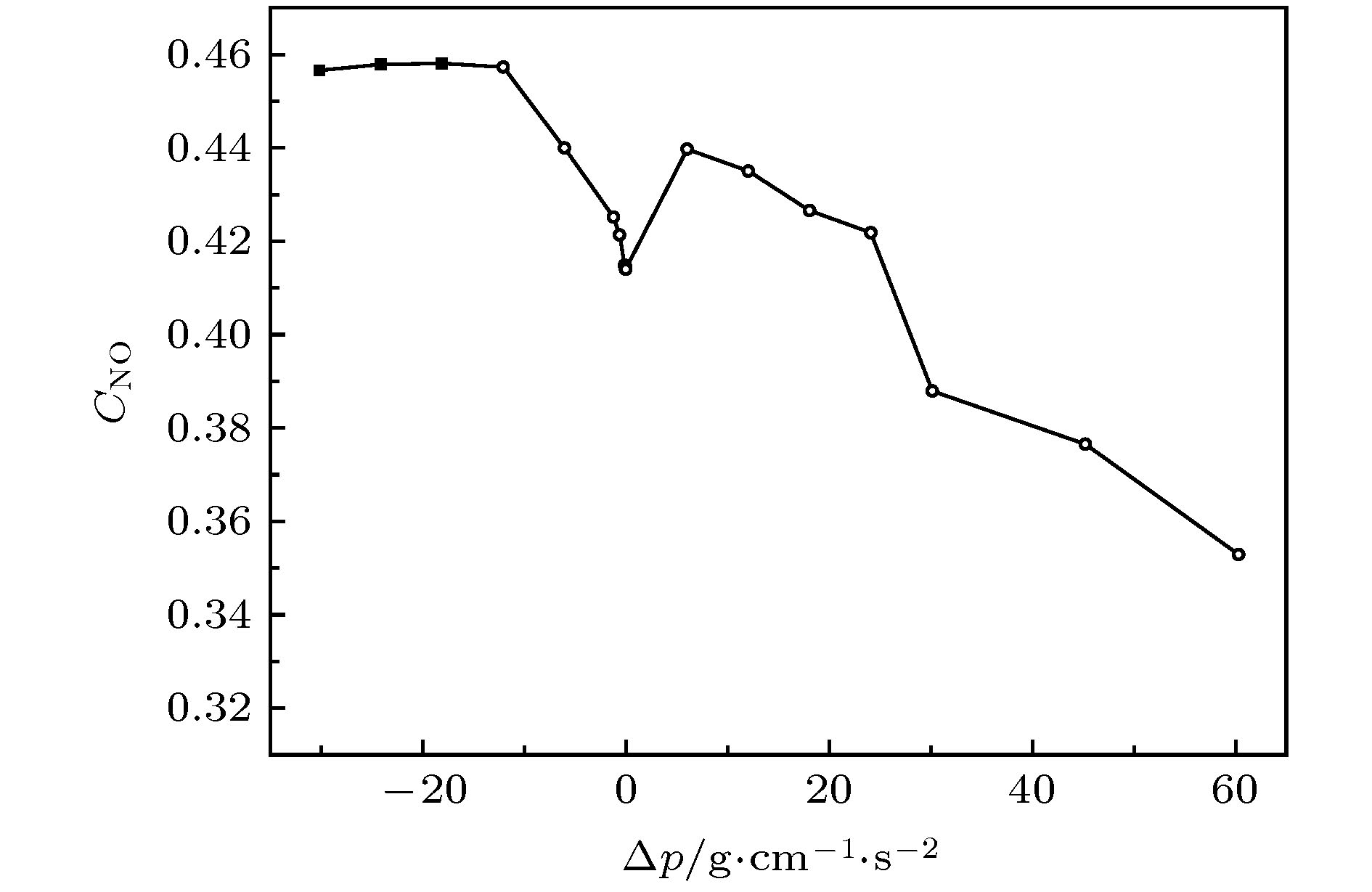
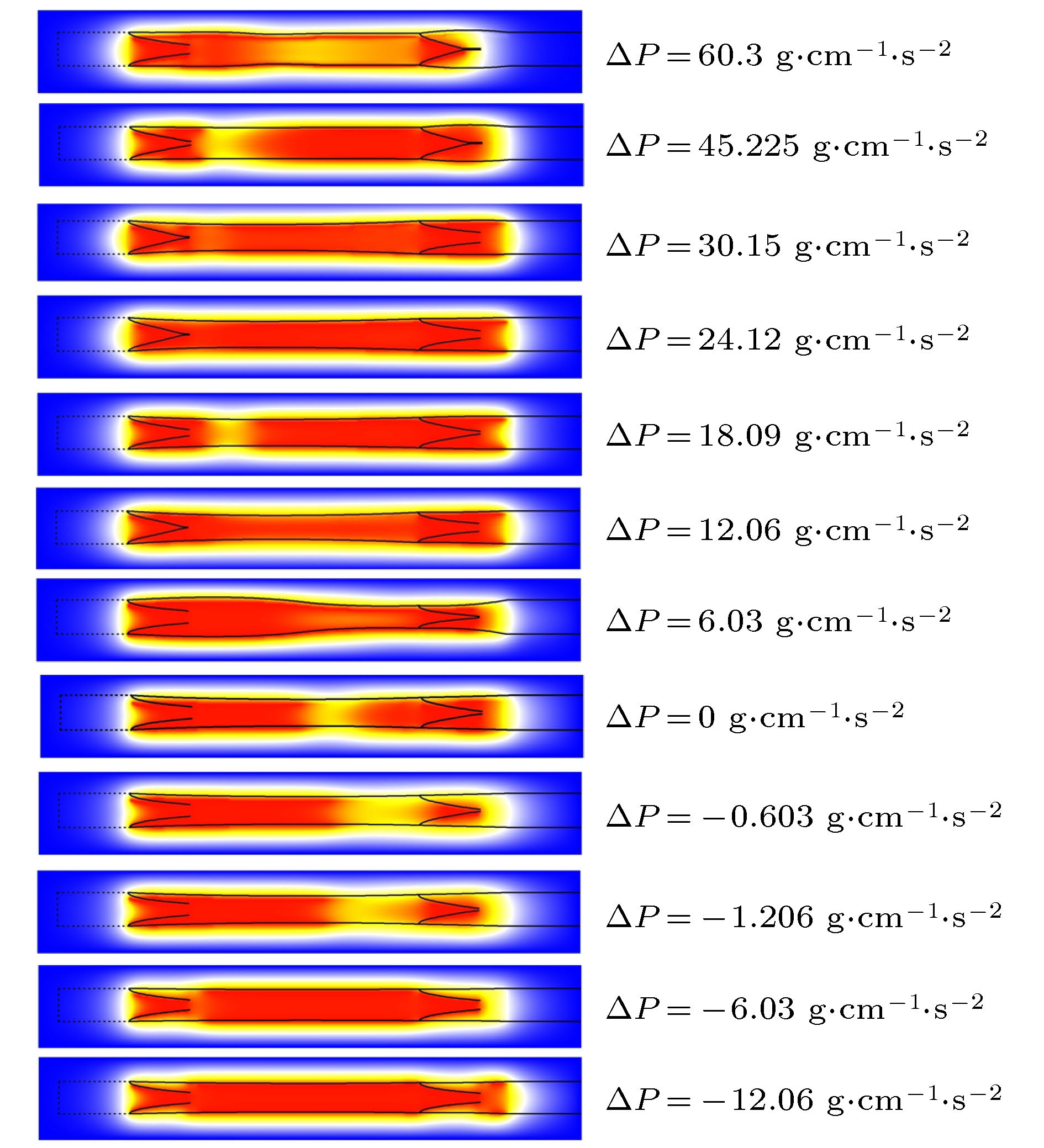
图 6 NO平均浓度与压强差关系图
Fig. 6. Relationship between NO average concentration and pressure difference.
表 1 Ca2+与NO的化学参数
Table 1. Chemical parameters of Ca2+ and NO.
参数 单位 数值 NO ${D_{{\rm{NO}}}}$ cm2/s 1.2 × 10–4 $K_{{\rm{NO}}}^ - $ s–1 3.7594 $K_{{\rm{NO}}}^ + $ 无量纲 400 Ca2+ ${D_{{\rm{Ca}}}}$ cm2/s 6.5 × 10–6 $K_{{\rm{Ca}}}^{-}$ s–1 37.6 $K_{{\rm{Ca}}}^{+}$ s–1 1.2 $K_\delta ^ + $ s–1 15038 ${C_{{\rm{th}}}}$ 无量纲 0.015 ${R_{{\rm{Ca}}}}$ cm 0.005 ${K_{{\rm{Ca}}, {\rm{NO}}}}$ 无量纲 5.3 h 无量纲 0.03  下载: 导出CSV
下载: 导出CSV
表 2 淋巴管与瓣膜参数
Table 2. Parameters of Lymphatic and valve.
参数 单位 数值 淋巴管 ${k_{\rm{M}}}$ ${\rm{dynes}}$ 7.6 × 10–5 ${k_{\rm{E}}}$ ${\rm{dynes}}/{{\rm{cm}}^{\rm{2}}}$ 4.52 ${k_{\rm{B}}}$ ${\rm{dynes}} /{{\rm{cm}}^2}$ 9045 ${k_{\rm{r}}}$ dynes·s/cm 4.8 × 10–9 ${k_{{\rm{NO}}}}$ 无量纲 1 ${R_{\rm{l}}}$ cm 0.003 ${R_{\rm{0}}}$ cm 0.005 瓣膜 $k_{\rm{B}}^\nu $ dynes /cm2 0—0.2 $k_{\rm{E}}^\nu $ dynes /cm2 9.0 × 10–4 $k_{\rm{r}}^\nu $ dynes /cm2 0.0091 A cm–1 1500 $\varDelta $ cm 2 × 10–4
下载: 导出CSV
表 3 出口压强高于入口压强时正压力差
Table 3. Positive pressure when outlet pressure is higher than inlet pressure.
正压力差 ${{\rho _{{\rm{out}}}}} /$g·cm–3 1.0020 1.0015 1.0010 1.0008 1.0006 1.0004 1.0002 ${\Delta \rho }/$g·cm–3 0.0020 0.0015 0.0010 0.0008 0.0006 0.0004 0.0002 ${\Delta P} /$g·cm–1·s–2 60.3 45.225 30.15 24.12 18.09 12.06 6.03
下载: 导出CSV
表 4 出口压强低于入口压强时负压力差
Table 4. Negative pressure when outlet pressure is lower than inlet pressure.
负压力差 ${\rho _{{\rm{out}}}}/$g·cm–3 1.0000 0.99998 0.99996 0.9998 0.9996 $\Delta \rho /$g·cm–3 0 –0.00002 –0.00004 –0.0002 –0.0004 $\Delta P/$g·cm–1·s–2 0 –0.603 –1.206 –6.03 –12.06
下载: 导出CSV
-
[1] Louveau A, Smirnov I, Keyes T J, Eccles J D, Rouhani S J, Peske J D, Derecki N C, Castle D, Mandell J W, Lee K S, Harris T H, Kipnis J 2015 Nature 523 377
[2] Margaris K N, Black R A 2012 J. R. Soc. Interface 9 601
Google Scholar
[3] Macdonald A J, Arkill K P, Tabor G R, McHale N G, Winlove C P 2008 Am. J. Physiol. Heart C. 295 305
Google Scholar
[4] 张立民 2012 ATP敏感性钾通道在一氧化氮调节失血性休克大鼠离体淋巴管泵功能中的作用 (张家口: 河北北方学院)
Zhang L M 2012 Role of ATP-Sensitive Potassium Channels in Nitric Oxide in Regulating the Function of Isolated Lymphatic Pump in Hemorrhagic Shock(Zhangjiakou: Hebei North University) (in Chinese)
[5] 秦立鹏, 牛春雨, 赵自刚 2011 生理科学进展 42 237
Qin L P, Niu C Y, Zhao Z G 2011 Advances in Physiological Sciences 42 237
[6] Kunert C, Baish J W, Liao S, Padera T P, Munn L L 2015 PNAS 112 10938
Google Scholar
[7] Baish J W, Kunert C, Padera T P, Munn LL 2016 PLoS Comput. Biol. 12 1005
[8] 赵彤彤 2018 多孔介质含天然气水合物多相流动LBM模拟 (太原: 太原理工大学)
Zhao T T 2018 LBM Simulation of Multiphase Flow of Natural Gas Hydrate in Porous Media (Taiyuan: Taiyuan University of Technology) (in Chinese)
[9] Shan X, Chen H 1993 Phys. Rev. E 47 1815
Google Scholar
[10] Li H B, Mei Y M, Maimon N, Padera T P, Baish J W, Munn L L 2019 SCIENTIFICREPORTS 9 2045
[11] Chen, Chen, Martnez, Matthaeus 1991 Phys. Rev. Lett. 67 27
[12] Qian Y H, D’HumièresD, Lallemand P 1992 Europhys. Lett. 17 479
Google Scholar
[13] Sukop M C, ThorneJr D T2010 Lattice Boltzmann Modeling: An Introduction for Geoscientists and Engineers (Berlin: Springer Publishing Company) p36
[14] Pujol F, Hodgson T, Martinezcorral I, Prats A C, Devenport D, Takeichi M, Genot E, Mäkinen T, Francis-West P, Garmy-Susini B, Tatin F 2017 Arterioscl. Thromb. Vas. Biol. 37 1732
Google Scholar
[15] Scallan J P, Davis M J 2013 J. Physiol. 591 250
[16] Kawai Y, Yokoyama Y, Kaidoh M 2010 Am. J. Physiol. 298 647
Google Scholar
[17] Ladd A J C, Verberg R 2001 J. Stat. Phys. 104 1191
Google Scholar
[18] He X, Doolen G 1997 J. Comput. Phys. 134 306
Google Scholar
[19] H Glenn B, Olga Yu G, Zawieja D C 2011 Ame. J. Physiol. Heart C. 301 1897
Google Scholar

 下载:
下载:
计量
- 文章访问数: 8361
- PDF下载量: 64
- 被引次数: 0
