










-
荧光寿命显微成像技术(fluorescence lifetime imaging microscopy, FLIM)具有特异性强、灵敏度高、可定量测量等优点, 被广泛应用于生物医学、材料学等领域的研究. 为使FLIM技术更好地适用于高通量数据的快速分析, 近年来涌现出多种荧光寿命分析的新算法. 其中, 相量分析法(phasor analysis, PA)通过将时间域的拟合转化为频率域的直接计算来获得荧光寿命值, 与传统的最小二乘拟合法相比, 不仅更加简便快速, 适用于低光子数情形, 而且便于使数据内容可视化和对数据进行聚类分析, 因此越来越受到科研人员的青睐. 本文详细阐述了相量分析法的基本原理及运用方法, 并在此基础上介绍了该方法在细胞代谢状态测量、蛋白质相互作用研究、细胞微环境测量, 以及辅助病理诊断和提高超分辨成像分辨率等方面的应用, 着重讨论了PA法在这些FLIM应用实例中的优势所在, 为相关领域的研究提供有益的参考. 最后, 对荧光寿命数据的相量分析及其应用的发展方向进行展望.Fluorescence lifetime imaging microscopy (FLIM) is widely used in biomedical, materials and other fields. It not only has strong specificity and high sensitivity, but also has the capability of quantitative measurement because the fluorescence lifetime is not affected by the intensity of excitation, the concentration of fluorophores and photobleaching, and consequently is able to monitor the changes of microenvironment and reflecting the interaction between molecules. However, its application is limited to some extent by the complexity of data analysis. In order to make FLIM technology more suitable for fast analysis of high-throughput data, a variety of new algorithms for fluorescence lifetime analysis have emerged in recent years, such as phasor analysis, maximum likelihood estimation, first-order moment, Bayesian analysis, and compressed sensing. Among them, the phasor analysis (PA) method obtains the fluorescence lifetime by converting the fitting in the time domain to the direct calculation in the frequency domain. Compared with traditional least-square fitting method, it is not only simpler and faster, but also more suitable for the case of low photon counts. In addition, in the PA approach to FLIM, the fluorescence decay is directly converted into a phasor diagram by simple mathematics, where the phasor points originating from different pixels in the image are represented by the positions in the phasor plot, and thus the graphical representation obtained by PA method is convenient for data visualization and cluster analysis. Therefore, it has become a simple and powerful analysis method for FLIM, and is increasingly favored by researchers. In this paper, the basic principle of PA method and how we can use it are described in detail. And on this basis, the latest application research progress of the method in cell metabolism state measurement, protein interaction study, cell microenvironment measurement, auxiliary pathological diagnosis, and resolution improvement in super-resolution imaging are introduced and summarized. The advantages of PA method in these FLIM applications are focused on, providing useful reference for the research in related fields. Finally, the phasor analysis method for FLIM data analysis and the development trend of its application are prospected.
-
Keywords:
- fluorescence lifetime microscopy /
- data analysis /
- phasor analysis /
- biomedical application
[1] 刘雄波, 林丹樱, 吴茜茜, 严伟, 罗腾, 杨志刚, 屈军乐 2018 物理学报 67 178701
 Google Scholar
Google Scholar
Liu X B, Lin D Y, Wu Q Q, Yan W, Luo T, Yang Z G, Qu J L 2018 Acta Phys. Sin. 67 178701
Google Scholar
[2] Levchenko S M, Pliss A, Qu J 2018 J. Innovative Opt. Health Sci. 11 1730009
Google Scholar
[3] 刘超, 周燕, 王新伟, 刘育梁 2011 激光与光电子学进展 48 111102
Google Scholar
Liu C, Zhou Y, Wang X, Liu Y 2011 Laser Optoelectron. Prog. 48 111102
Google Scholar
[4] Fitzgerald C, Hosny N A, Tong H, Seville P C, Gallimore P J, Davidson N M, Athanasiadis A, Botchway S W, Ward A D, Kalberer M, Kuimov M K, Pope F D 2016 Phys. Chem. Chem. Phys. 18 21710
Google Scholar
[5] Gratton E, Breusegem S, Sutin J, Ruan Q, Barry N 2003 J. Biomed. Opt. 8 38190
[6] Suman R, Leonel M, Jameson D M, Gratton E 2018 Nat. Protoc. 13 1979
Google Scholar
[7] Chessel A, Waharte F, Salamero J, Kervrann C 2013 21st European Signal Processing Conference Marrakech, Morocco, September 9–13, 2013 p1
[8] 徐玲玲 2013 博士学位论文 (武汉: 华中科技大学)
Xu L L 2013 Ph. D. Dissertation (Wuhan: Huazhong University of Science and Technology) (in Chinese)
[9] Rowley M I, Coolen A C C, Vojnovic B, Barber P R 2016 PLoS One 11 e0158404
Google Scholar
[10] Yang S, Lee J, Lee Y, Lee M, Lee B U 2015 J. Biomed. Opt. 20 096003
Google Scholar
[11] Liu X, Lin D, Becker W, Niu J, Yu B, Liu L, Qu J 2019 J. Innovative. Opt. Health Sci. 12 1930003
Google Scholar
[12] Stefl M, James N G, Ross J A, Jameson D M 2011 Anal. Biochem. 410 62
Google Scholar
[13] Jameson D M, Gratton E, Hall R D 1984 Appl. Spectrosc. Rev. 20 55
Google Scholar
[14] Weber G 1981 J. Phys. Chem. B 85 949
Google Scholar
[15] Redford G I, Clegg R M 2005 J. Fluoresc. 15 805
Google Scholar
[16] Digman M A, Caiolfa V R, Zamai M, Gratton E 2008 Biophys. J. 94 L14
Google Scholar
[17] Bird D K, Yan L, Vrotsos K M, Eliceiri K W, Vaughan E M, Keely P J, White J G, Ramanujam N 2005 Cancer Res. 65 8766
Google Scholar
[18] Stringari C, Cinquin A, Cinquin O, Digman M A, Donovan P J, Gratton E 2011 Proc. Natl. Acad. Sci. U.S.A. 108 13582
Google Scholar
[19] Stringari C, Edwards R A, Pate K T, Waterman M L, Donovan P J, Gratton E 2012 Sci. Rep. 2 568
Google Scholar
[20] Stringari C, Donovan P, Gratton E 2012 Proc. SPIE San Francisco, CA January 22–24, 2012 p9
[21] Lee D H, Li X, Ma N, Digman M A, Lee A P 2018 Lab Chip 18 1349
Google Scholar
[22] Romero-López M, Trinh A L, Sobrino A, Hatch M M S, Keating M T, Fimbres C, Lewis D E, Gershon P D, Botvinick E L, Digman M, Lowengrub J S, Hughes C C W 2016 Biomaterials 116 118
[23] Sameni S, Syed A, Marsh J L, Digman M A 2016 Sci. Rep. 6 34755
Google Scholar
[24] Dong Y, Sameni1 S, Digman M A, Brewer G J 2019 Sci. Rep. 9 11274
Google Scholar
[25] Dong Y, Digman M A, Brewer G J 2019 GeroScience 41 51
Google Scholar
[26] Hato T, Winfree S, Day R, Sandoval R M, Molitoris B A, Yoder M C, Wiggins R C, Zheng Y, Dunn K W, Dagher P C 2017 J. Am. Soc. Nephrol. 28 2420
Google Scholar
[27] Datta R, Heylman C, George S C, Gratton E 2016 Biomed. Opt. Express 7 1690
Google Scholar
[28] Hinde E, Digman M A, Hahn K M, Hahn K M, Gratton E 2012 Microsc. Res. Tech. 75 271
Google Scholar
[29] Hinde E, Digman M A, Hahn K M, Gratton E 2013 Proc. Natl. Acad. Sci. U.S.A. 110 135
Google Scholar
[30] Lou J Q, Scipioni L, Wright B K, Bartolec T K, Zhang J, Masamsetti V P, Gaus K, Gratton E, Cesare A J, Hinde E 2019 Proc. Natl. Acad. Sci. U.S.A. 116 7323
Google Scholar
[31] Chen H, Ma N, Kagawa K, Kawahito S, Digman M, Gratton E 2018 J. Biophotonics 12 e201800223
[32] Battisti A, Digman M A, Gratton E, Storti B, Beltram F, Bizzarri R 2012 Chem. Commun. 48 5127
Google Scholar
[33] Zhou T, Luo T, Song J, Qu J 2018 Anal. Chem. 90 2170
Google Scholar
[34] Ferri G, Nucara L, Biver T, Battisti A, Signore G, Bizzarri R 2016 Biophys. J. 110 163a
Google Scholar
[35] Luo T, Lu Y, Liu S, Lin D, Qu J 2017 Anal. Chem. 89 8104
Google Scholar
[36] Luo T, Lu Y, Liu S, Lin D, Qu J 2017 Anal. Chem. 89 9224
Google Scholar
[37] Ranjit S, Dvornikov A, Levi M, Furgeson S, Gratton E 2016 Biomed. Opt. Express 7 3519
Google Scholar
[38] Lanzanò L, Hernandez I C, Castello M, Gratton E, Diaspro A, Vicidomini G 2015 Nat. Commun. 6 6701
Google Scholar
[39] Wang L, Chen B, Yan W, Yang Z, Peng X, Lin D, Weng X, Ye T, Qu J 2018 Nanoscale 10 16252
Google Scholar
[40] Tortarolo G, Sun Y, Teng K W, Ishitsuka Y, Lanzano L, Selvin P R, Barbieri B, Diaspro A, Vicidomini G 2019 Nanoscale 11 1754
Google Scholar
[41] 周宇会, 魏九峰, 李国东, 刘明 2017 国际肿瘤学杂志 44 762
Google Scholar
Zhou Y, Wei J, Li G, Liu M 2017 J. Int. Oco. 44 762
Google Scholar
[42] 林丹樱, 屈军乐 2017 物理学报 66 148703
Google Scholar
Lin D Y, Qu J L 2017 Acta Phys. Sin. 66 148703
Google Scholar
-
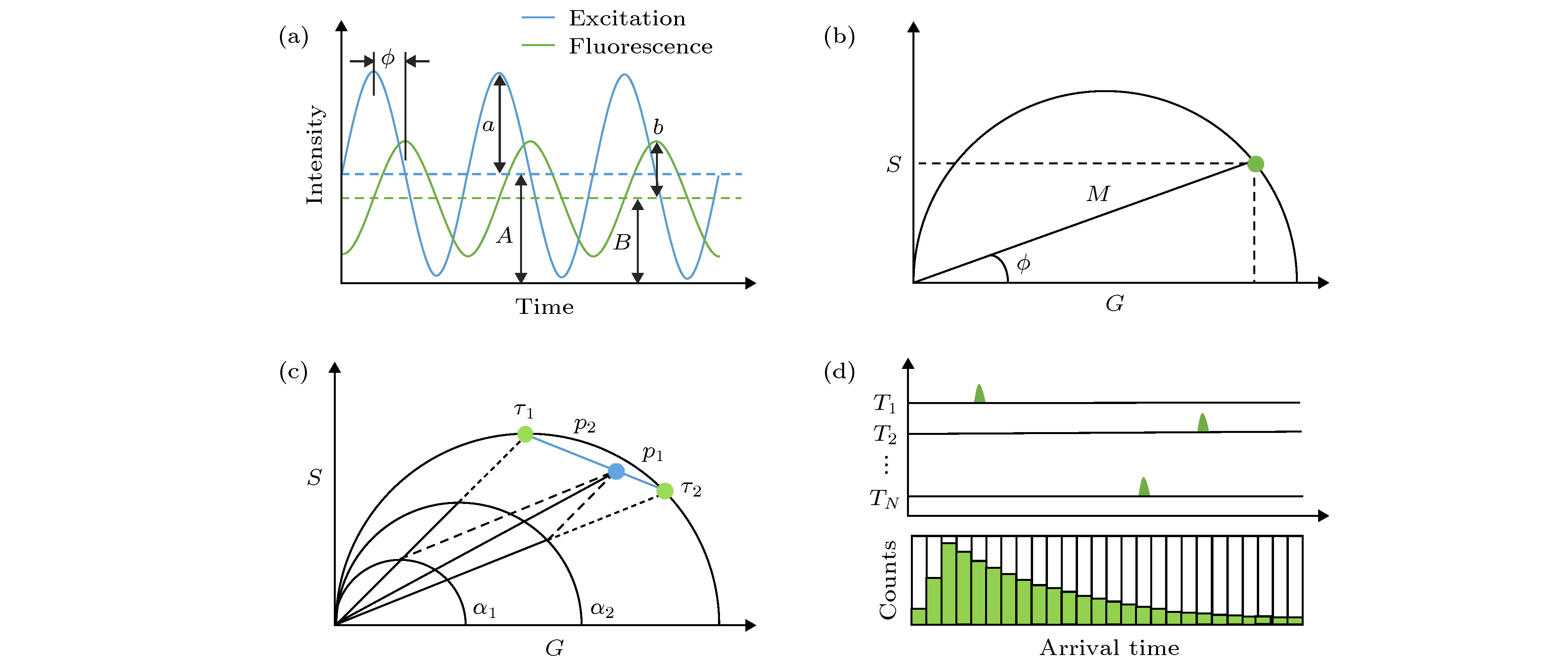
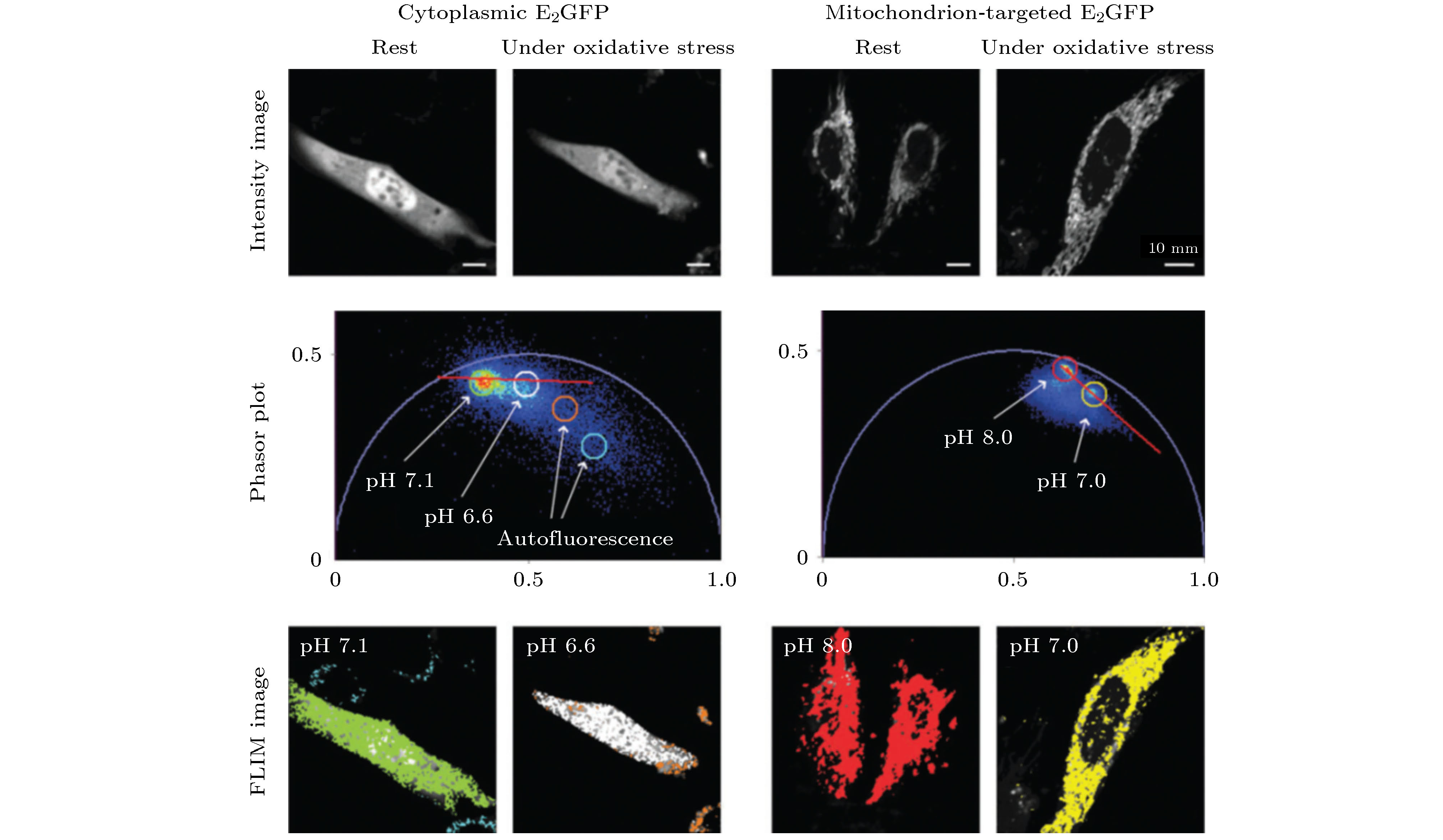
图 1 荧光寿命的测量方法及相量分析(PA)法示意图 (a)频域法测量原理示意图; (b)单指数衰减的寿命相量示例图; (c)双指数衰减的寿命相量示例图; (d)时间相关单光子计数(TCSPC)测量原理示意图
Fig. 1. Schematic diagram of fluorescence lifetime measurement and phasor analysis (PA):(a) Frequency domain method; (b) lifetime phasor of single-exponential decay; (c) lifetime phasor of bi-exponential decay; (d) time-correlated single photon counting (TCSPC) method.
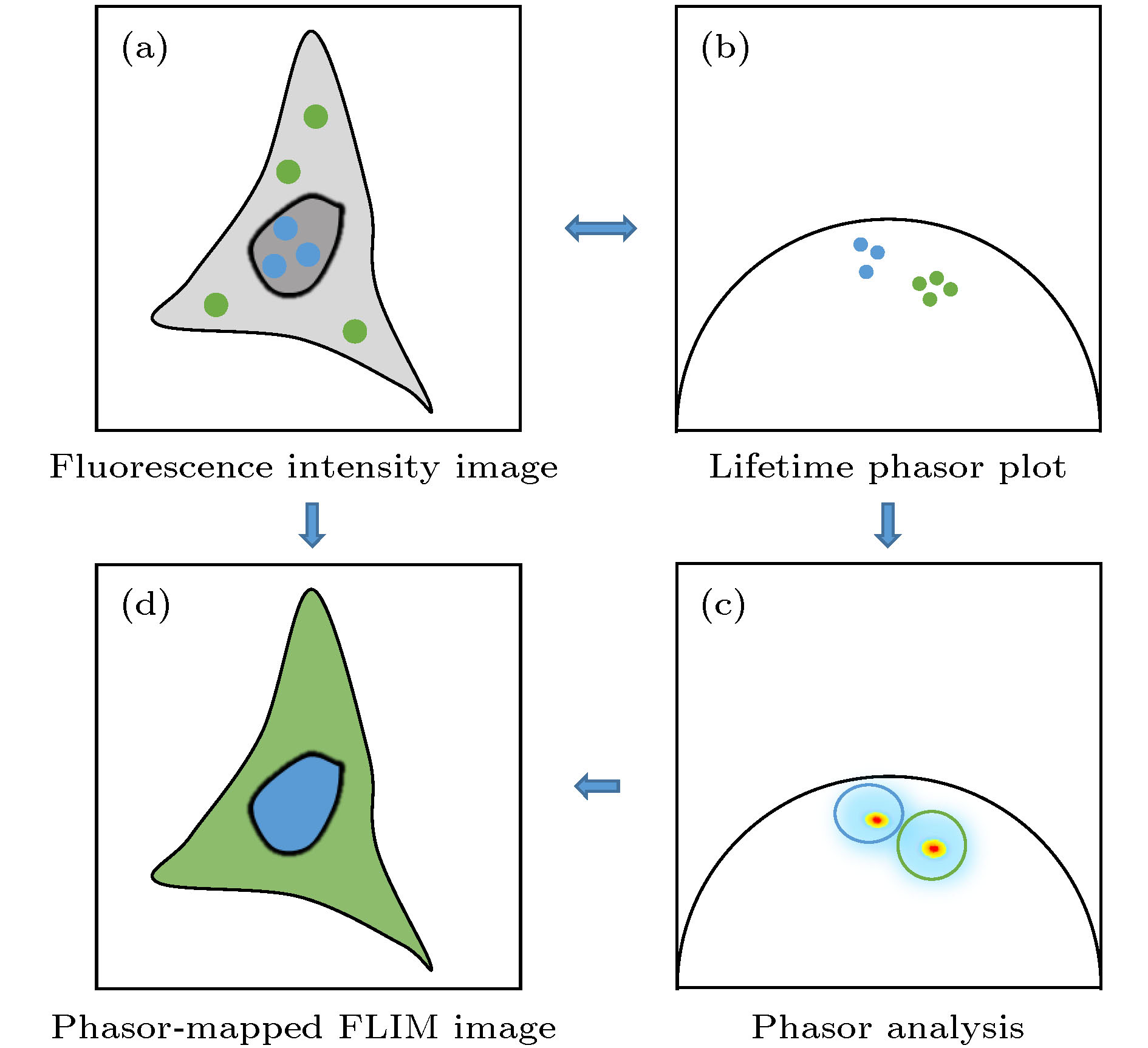
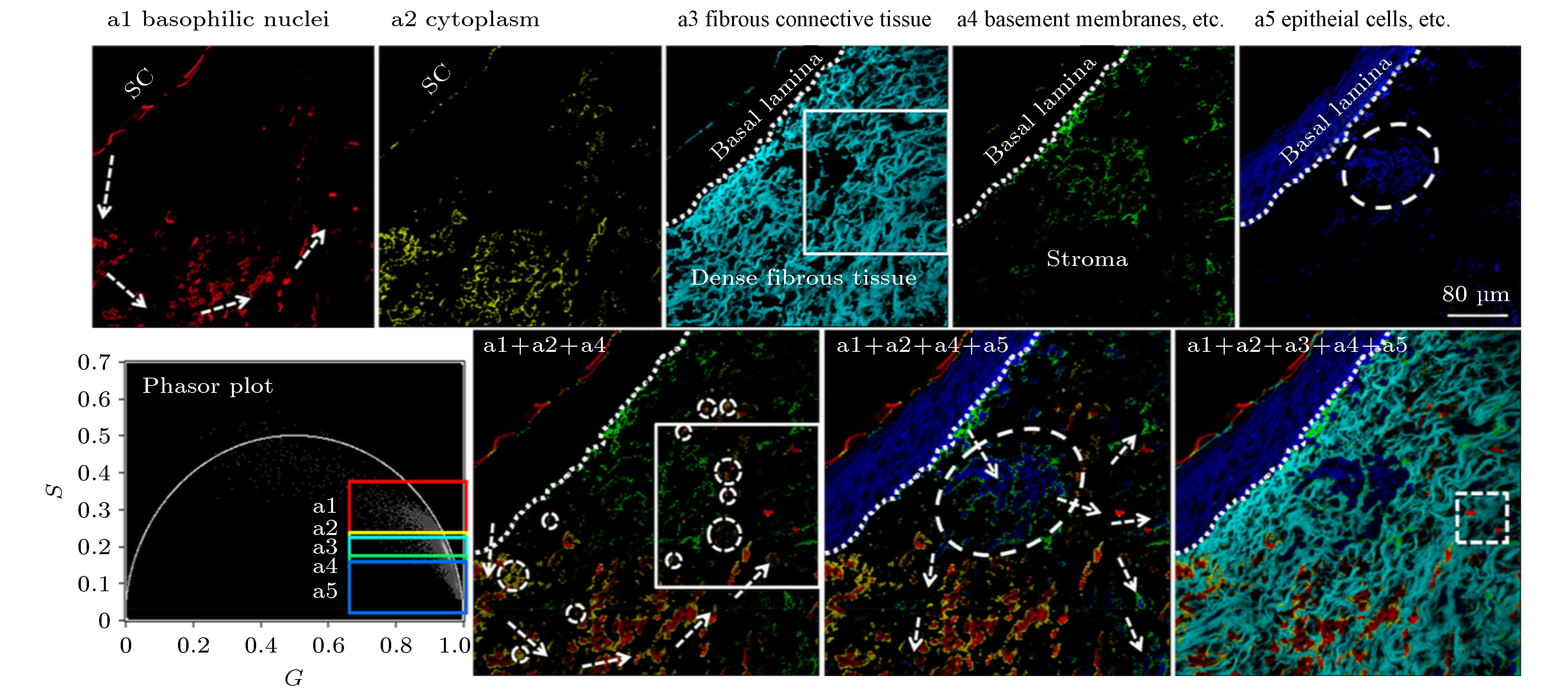
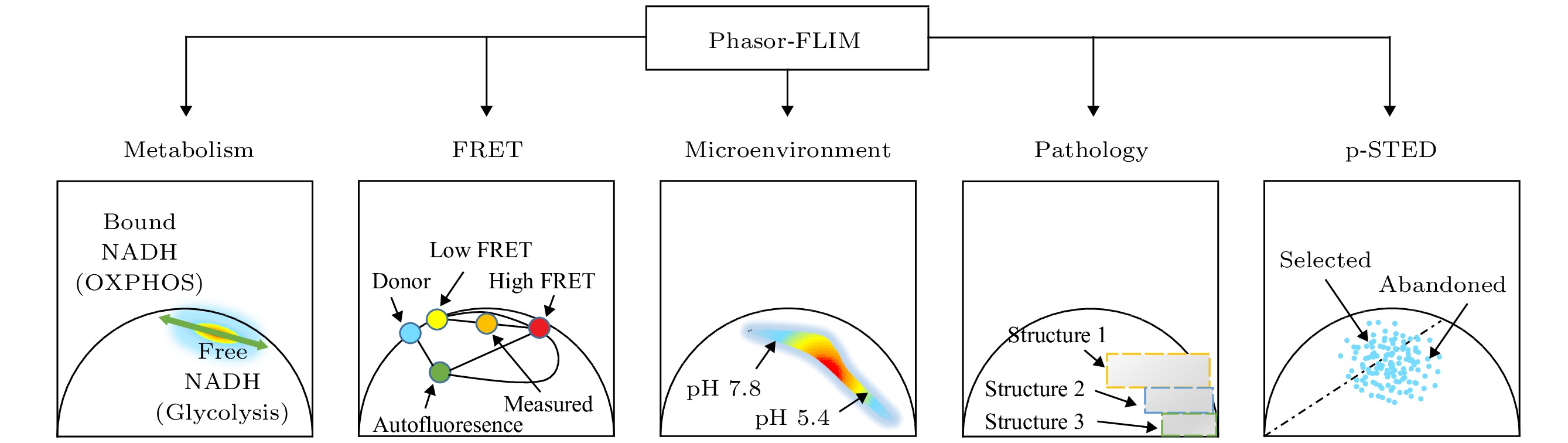
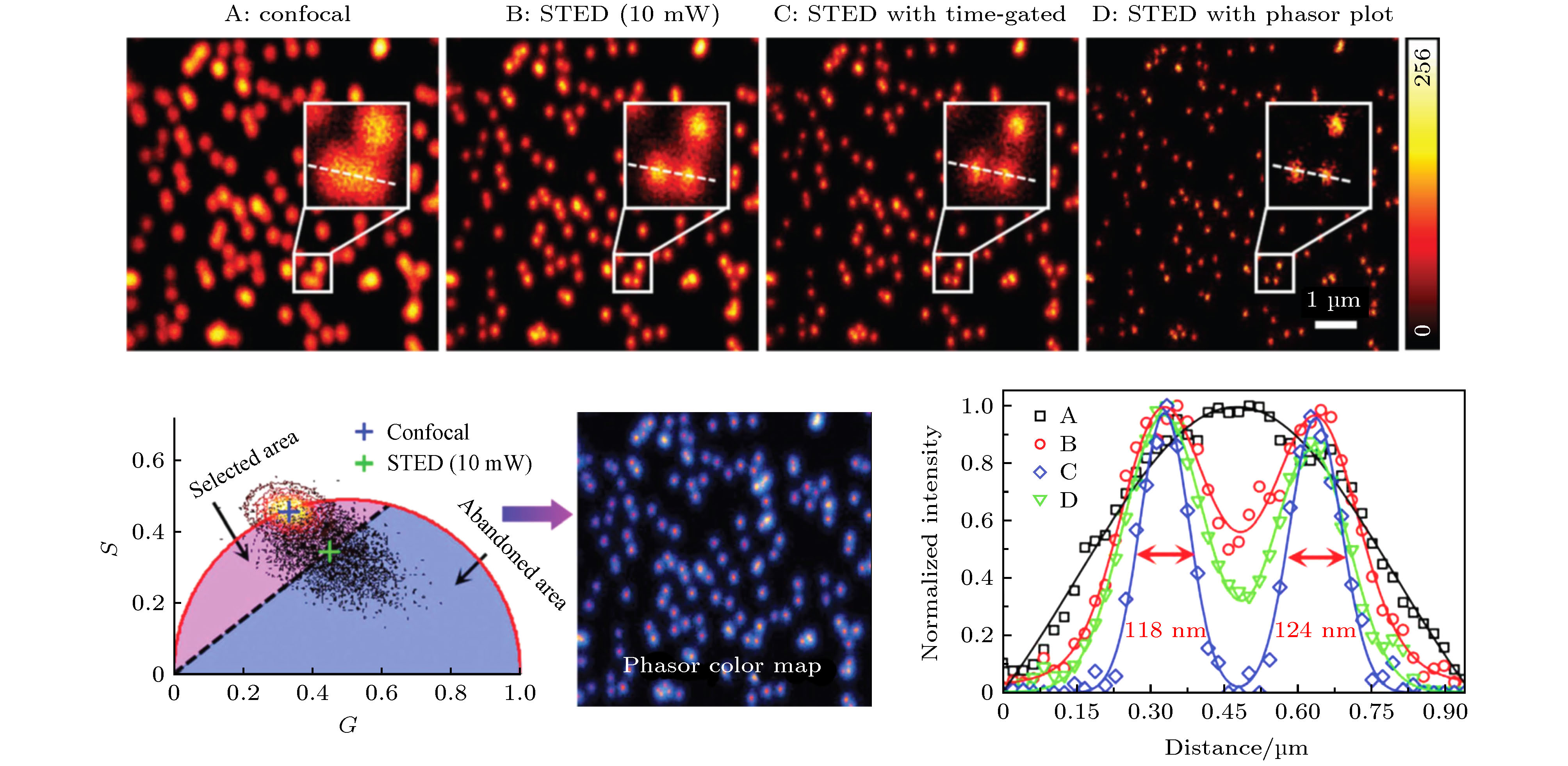
图 2 Phasor-FLIM的应用思路示意图 (a)包含未处理寿命信息的荧光强度图; (b)经PA法分析得到的寿命相量图; (c)对寿命相量直接进行分析; (d)通过相量聚类分析和伪彩色标记得到的荧光寿命图
Fig. 2. Schematic diagram of phasor-FLIM application:(a) Fluorescence intensity image with untreated lifetime information; (b) lifetime phasor plot obtained by PA analysis; (c) direct analysis of lifetime phasors; (d) phasor-mapped FLIM image based on phasor clustering analysis and pseudo-color assignment.
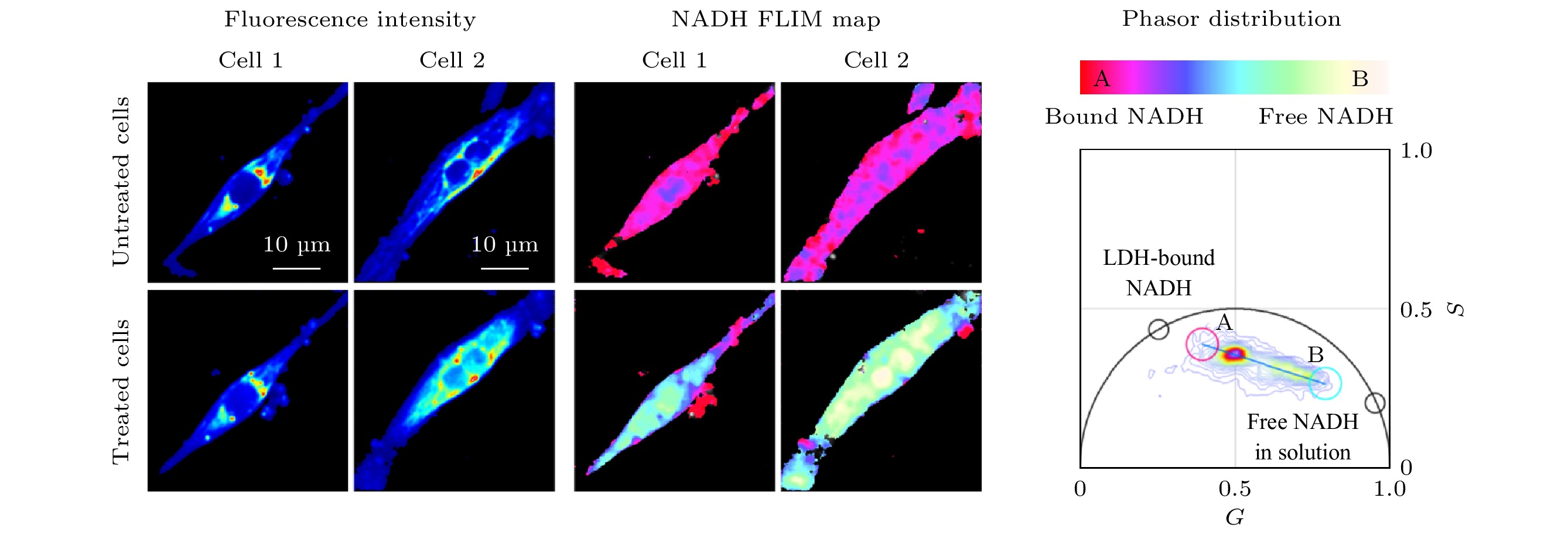
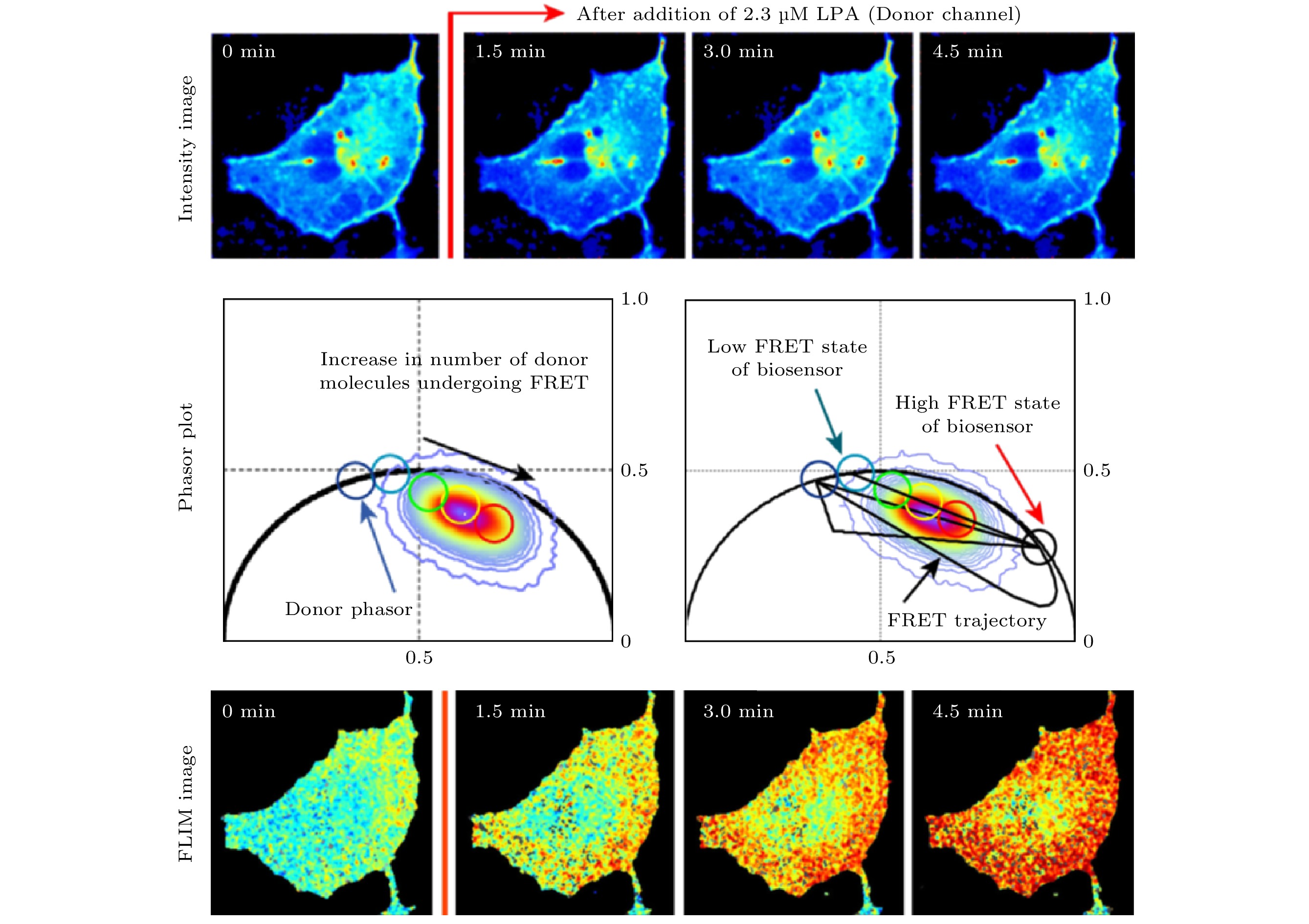
-
[1] 刘雄波, 林丹樱, 吴茜茜, 严伟, 罗腾, 杨志刚, 屈军乐 2018 物理学报 67 178701
Google Scholar
Liu X B, Lin D Y, Wu Q Q, Yan W, Luo T, Yang Z G, Qu J L 2018 Acta Phys. Sin. 67 178701
Google Scholar
[2] Levchenko S M, Pliss A, Qu J 2018 J. Innovative Opt. Health Sci. 11 1730009
Google Scholar
[3] 刘超, 周燕, 王新伟, 刘育梁 2011 激光与光电子学进展 48 111102
Google Scholar
Liu C, Zhou Y, Wang X, Liu Y 2011 Laser Optoelectron. Prog. 48 111102
Google Scholar
[4] Fitzgerald C, Hosny N A, Tong H, Seville P C, Gallimore P J, Davidson N M, Athanasiadis A, Botchway S W, Ward A D, Kalberer M, Kuimov M K, Pope F D 2016 Phys. Chem. Chem. Phys. 18 21710
Google Scholar
[5] Gratton E, Breusegem S, Sutin J, Ruan Q, Barry N 2003 J. Biomed. Opt. 8 38190
[6] Suman R, Leonel M, Jameson D M, Gratton E 2018 Nat. Protoc. 13 1979
Google Scholar
[7] Chessel A, Waharte F, Salamero J, Kervrann C 2013 21st European Signal Processing Conference Marrakech, Morocco, September 9–13, 2013 p1
[8] 徐玲玲 2013 博士学位论文 (武汉: 华中科技大学)
Xu L L 2013 Ph. D. Dissertation (Wuhan: Huazhong University of Science and Technology) (in Chinese)
[9] Rowley M I, Coolen A C C, Vojnovic B, Barber P R 2016 PLoS One 11 e0158404
Google Scholar
[10] Yang S, Lee J, Lee Y, Lee M, Lee B U 2015 J. Biomed. Opt. 20 096003
Google Scholar
[11] Liu X, Lin D, Becker W, Niu J, Yu B, Liu L, Qu J 2019 J. Innovative. Opt. Health Sci. 12 1930003
Google Scholar
[12] Stefl M, James N G, Ross J A, Jameson D M 2011 Anal. Biochem. 410 62
Google Scholar
[13] Jameson D M, Gratton E, Hall R D 1984 Appl. Spectrosc. Rev. 20 55
Google Scholar
[14] Weber G 1981 J. Phys. Chem. B 85 949
Google Scholar
[15] Redford G I, Clegg R M 2005 J. Fluoresc. 15 805
Google Scholar
[16] Digman M A, Caiolfa V R, Zamai M, Gratton E 2008 Biophys. J. 94 L14
Google Scholar
[17] Bird D K, Yan L, Vrotsos K M, Eliceiri K W, Vaughan E M, Keely P J, White J G, Ramanujam N 2005 Cancer Res. 65 8766
Google Scholar
[18] Stringari C, Cinquin A, Cinquin O, Digman M A, Donovan P J, Gratton E 2011 Proc. Natl. Acad. Sci. U.S.A. 108 13582
Google Scholar
[19] Stringari C, Edwards R A, Pate K T, Waterman M L, Donovan P J, Gratton E 2012 Sci. Rep. 2 568
Google Scholar
[20] Stringari C, Donovan P, Gratton E 2012 Proc. SPIE San Francisco, CA January 22–24, 2012 p9
[21] Lee D H, Li X, Ma N, Digman M A, Lee A P 2018 Lab Chip 18 1349
Google Scholar
[22] Romero-López M, Trinh A L, Sobrino A, Hatch M M S, Keating M T, Fimbres C, Lewis D E, Gershon P D, Botvinick E L, Digman M, Lowengrub J S, Hughes C C W 2016 Biomaterials 116 118
[23] Sameni S, Syed A, Marsh J L, Digman M A 2016 Sci. Rep. 6 34755
Google Scholar
[24] Dong Y, Sameni1 S, Digman M A, Brewer G J 2019 Sci. Rep. 9 11274
Google Scholar
[25] Dong Y, Digman M A, Brewer G J 2019 GeroScience 41 51
Google Scholar
[26] Hato T, Winfree S, Day R, Sandoval R M, Molitoris B A, Yoder M C, Wiggins R C, Zheng Y, Dunn K W, Dagher P C 2017 J. Am. Soc. Nephrol. 28 2420
Google Scholar
[27] Datta R, Heylman C, George S C, Gratton E 2016 Biomed. Opt. Express 7 1690
Google Scholar
[28] Hinde E, Digman M A, Hahn K M, Hahn K M, Gratton E 2012 Microsc. Res. Tech. 75 271
Google Scholar
[29] Hinde E, Digman M A, Hahn K M, Gratton E 2013 Proc. Natl. Acad. Sci. U.S.A. 110 135
Google Scholar
[30] Lou J Q, Scipioni L, Wright B K, Bartolec T K, Zhang J, Masamsetti V P, Gaus K, Gratton E, Cesare A J, Hinde E 2019 Proc. Natl. Acad. Sci. U.S.A. 116 7323
Google Scholar
[31] Chen H, Ma N, Kagawa K, Kawahito S, Digman M, Gratton E 2018 J. Biophotonics 12 e201800223
[32] Battisti A, Digman M A, Gratton E, Storti B, Beltram F, Bizzarri R 2012 Chem. Commun. 48 5127
Google Scholar
[33] Zhou T, Luo T, Song J, Qu J 2018 Anal. Chem. 90 2170
Google Scholar
[34] Ferri G, Nucara L, Biver T, Battisti A, Signore G, Bizzarri R 2016 Biophys. J. 110 163a
Google Scholar
[35] Luo T, Lu Y, Liu S, Lin D, Qu J 2017 Anal. Chem. 89 8104
Google Scholar
[36] Luo T, Lu Y, Liu S, Lin D, Qu J 2017 Anal. Chem. 89 9224
Google Scholar
[37] Ranjit S, Dvornikov A, Levi M, Furgeson S, Gratton E 2016 Biomed. Opt. Express 7 3519
Google Scholar
[38] Lanzanò L, Hernandez I C, Castello M, Gratton E, Diaspro A, Vicidomini G 2015 Nat. Commun. 6 6701
Google Scholar
[39] Wang L, Chen B, Yan W, Yang Z, Peng X, Lin D, Weng X, Ye T, Qu J 2018 Nanoscale 10 16252
Google Scholar
[40] Tortarolo G, Sun Y, Teng K W, Ishitsuka Y, Lanzano L, Selvin P R, Barbieri B, Diaspro A, Vicidomini G 2019 Nanoscale 11 1754
Google Scholar
[41] 周宇会, 魏九峰, 李国东, 刘明 2017 国际肿瘤学杂志 44 762
Google Scholar
Zhou Y, Wei J, Li G, Liu M 2017 J. Int. Oco. 44 762
Google Scholar
[42] 林丹樱, 屈军乐 2017 物理学报 66 148703
Google Scholar
Lin D Y, Qu J L 2017 Acta Phys. Sin. 66 148703
Google Scholar

 下载:
下载:
计量
- 文章访问数: 20593
- PDF下载量: 398
- 被引次数: 0
