










-
神经元放电率自稳态是指大脑神经网络的放电率维持在相对稳定的状态. 大量实验研究发现神经元放电率自稳态是神经电活动的重要特征, 并且放电率自稳态是实现神经信息处理及维持正常脑功能的基础, 因此放电率自稳态的研究是神经科学领域的重要科学问题. 脑神经网络是一个高度复杂的动态系统, 存在大量输入扰动信号及由于动态链接导致的参数摄动, 因此如何建立并维持神经元放电率自稳态及其鲁棒性仍有待深入研究. 反馈神经回路是皮层神经网络的典型连接模式, 抑制性突触可塑性对神经元放电率自稳态具有重要的调控作用. 本文通过构建包含抑制性突触可塑性的反馈神经回路模型对神经元放电率自稳态及其鲁棒性进行计算研究. 结果表明: 在抑制性突触可塑性的作用下, 神经元放电率可自适应地跟踪目标放电率, 从而取得放电率自稳态; 在有外部输入干扰和参数摄动的情况下, 神经元放电率具有良好的抗扰动性能, 表明放电率自稳态具有很强的鲁棒性; 理论分析揭示了抑制性突触可塑性学习规则是神经元放电率自稳态的神经机制; 仿真分析进一步揭示了学习率及目标放电率对放电率自稳态建立过程具有重要影响.Neural firing rate homeostasis, as an important feature of neural electrical activity, means that the firing rate in brain is maintained in a relatively stable state, and fluctuates around a constant value. Extensive experimental studies have revealed that the firing rate homeostasis is ubiquitous in brain, and provides a base for neural information processing and maintaining normal neurological functions, so that the research on neural firing rate homeostasis is a central problem in the field of neuroscience. Cortical neural network is a highly complex dynamic system with a large number of input disturbance signals and parameter perturbations due to dynamic connection. However, it remains to be further investigated how firing rate homeostasis is established in cortical neural network, furthermore, maintains robustness to these disturbances and perturbations. The feedback neural circuit with recurrent excitatory and inhibitory connection is a typical connective pattern in cortical cortex, and inhibitory synaptic plasticity plays a crucial role in achieving neural firing rate homeostasis. Here, by constructing a feedback neural network with inhibitory spike timing-dependent plasticity (STDP), we conduct a computational research to elucidate the mechanism of neural firing rate homeostasis. The results indicate that the neuronal firing rate can track the target firing rate accurately under the regulation of inhibitory synaptic plasticity, thus achieve firing rate homeostasis. In the face of external disturbances and parameter perturbations, the neuron firing rate deviates transiently from the target firing rate value, and converges to the target firing rate value at a steady state, which demonstrates that the firing rate homeostasis established by the inhibitory synaptic plasticity can maintain strong robustness. Furthermore, the analytical research qualitatively explains the firing rate homeostasis mechanism underlined by inhibitory synaptic plasticity. Finally, the simulations further demonstrate that the learning rate value and the firing rate set point value also exert a quantitative influence on the firing rate homeostasis. Overall, these findings not only gain an insight into the firing rate homeostasis mechanism underlined by inhibitory synaptic plasticity, but also inspire testable hypotheses for future experimental studies.
-
Keywords:
- inhibitory synaptic plasticity /
- firing rate homeostasis /
- robustness
[1] Gläser C, Joublin F 2011 IEEE T. Auton. Ment. De. 3 285
 Google Scholar
Google Scholar
[2] Hengen K B, Lambo M E, Hooser S D, van Katz D B, Turrigiano G G 2013 Neuron 80 335
Google Scholar
[3] Corner M A, Ramakers G J A 1992 Dev. Brain Res. 65 57
Google Scholar
[4] Ramakers G J A, Corner M A, Habets A M M C 1990 Exp. Brain Res. 79 157
Google Scholar
[5] Ramakers G J A, Galen H V, Feenstra M G P, Corner M A, Boer G J 1994 Int. J. Dev. Neurosci. 12 611
Google Scholar
[6] Pol A N V D, Obrietan K, Belousov A 1996 Neuroscience 74 653
Google Scholar
[7] Turrigiano G G, Leslie K R, Desai N S, Rutherford L C, Nelson S B 1998 Nature 391 892
Google Scholar
[8] Rutherford L C, Nelson S B, Turrigiano G G 1998 Neuron 21 521
Google Scholar
[9] Burrone J, O'Byrne M, Murthy V N 2002 Nature 420 414
Google Scholar
[10] Turrigiano G G, Nelson S B 2004 Nat. Rev. Neurosci. 5 97
Google Scholar
[11] Turrigiano G 2012 CSH Perspect. Biol. 4 a005736
Google Scholar
[12] Cannon J, Miller P 2016 J. Neurophysiol. 116 2004
Google Scholar
[13] Cannon J, Miller P 2017 J. Math. Neurosc. 7 1
Google Scholar
[14] Miller P, Cannon J 2018 Biol. Cybern. 113 47
[15] McClelland J L, McNaughton B L, O'Reilly R C 1995 Psychol. Rev. 102 419
Google Scholar
[16] Frankland P W, O'Brien C, Ohno M, Kirkwood A, Silva A J 2001 Nature 411 309
Google Scholar
[17] Carcea I, Froemke R C 2013 Prog. Brain. Res. 207 65
Google Scholar
[18] Martin S J, Grimwood P D, Morris R G M 2000 Annu. Rev. Neurosci. 23 649
Google Scholar
[19] Sanderson J L, Dell'Acqua M L 2011 Neuroscientist 17 321
Google Scholar
[20] Yong L, Kauer J A 2010 Synapse 51 1
Google Scholar
[21] Haas J S, Thomas N, Abarbanel H D I 2006 J. Neurophysiol. 96 3305
Google Scholar
[22] D'Amour J A, Froemke R C 2015 Neuron 86 514
Google Scholar
[23] Hartmann K, Bruehl C, Golovko T, Draguhn A 2008 Plos One 3 e2979
Google Scholar
[24] Tohru K, Kazumasa Y, Yumiko Y, Crair M C, Yukio K 2008 Neuron 57 905
Google Scholar
[25] Stephen G, James R W 2001 Cereb. Cortex 11 37
Google Scholar
[26] Luz Y, Shamir M 2012 Plos. Comput. Biol. 8 e1002334
Google Scholar
[27] Hennequin G, Agnes E J, Vogels T P 2017 Annu. Revi. Neurosci. 40 557
Google Scholar
[28] Park H J, Friston K 2013 Science 342 1238411
Google Scholar
[29] Isaacson J S, Massimo S 2011 Neuronv 72 231
Google Scholar
[30] Maass W, Joshi P, Sontag E D 2007 Plos Comput. Biol. 3 e165
Google Scholar
[31] Jansen B H, Rit V G 1995 Biol. Cybern. 73 357
Google Scholar
[32] Chacron M J, André L, Leonard M 2005 Phys. Rev. E 72 051917
Google Scholar
[33] Froemke R C, Jones B J 2011 Neurosci. Biobehav. R. 35 2105
Google Scholar
[34] 王俊松, 徐瑶 2014 物理学报 63 068701
Google Scholar
Wang J S, Xu Y 2014 Acta Phys. Sin. 63 068701
Google Scholar
[35] 王美丽, 王俊松 2015 物理学报 64 108701
Google Scholar
Wang M L, Wang J S 2015 Acta Phys. Sin. 64 108701
Google Scholar
[36] Vogels T P, Abbott L F 2009 Nat. Neurosci. 12 483
Google Scholar
[37] Stepp N, Plenz D, Srinivasa N 2015 Plos Comput. Biol. 11 e1004043
Google Scholar
[38] Vogels T P, Sprekeler H, Zenke F, Clopath C, Gerstner W 2011 Science 334 1569
Google Scholar
[39] Maass W 2014 P. IEEE 102 860
Google Scholar
[40] Mcdonnell M D, Ward L M 2011 Nat. Rev. Neurosci. 12 183
Google Scholar
[41] Garrett D D, Mcintosh A R, Grady C L 2011 Nat. Rev. Neurosci. 12 612
Google Scholar
[42] Mcdonnell M D, Ward L M 2011 Nat. Rev. Neurosci. 12 415
Google Scholar
[43] Turrigiano G G 2008 Cell 135 422
Google Scholar
[44] Marder E, Tang L S 2010 Neuron 66 161
Google Scholar
[45] Sharon B, Dickman D K, Davis G W 2010 Neuron 66 220
Google Scholar
-
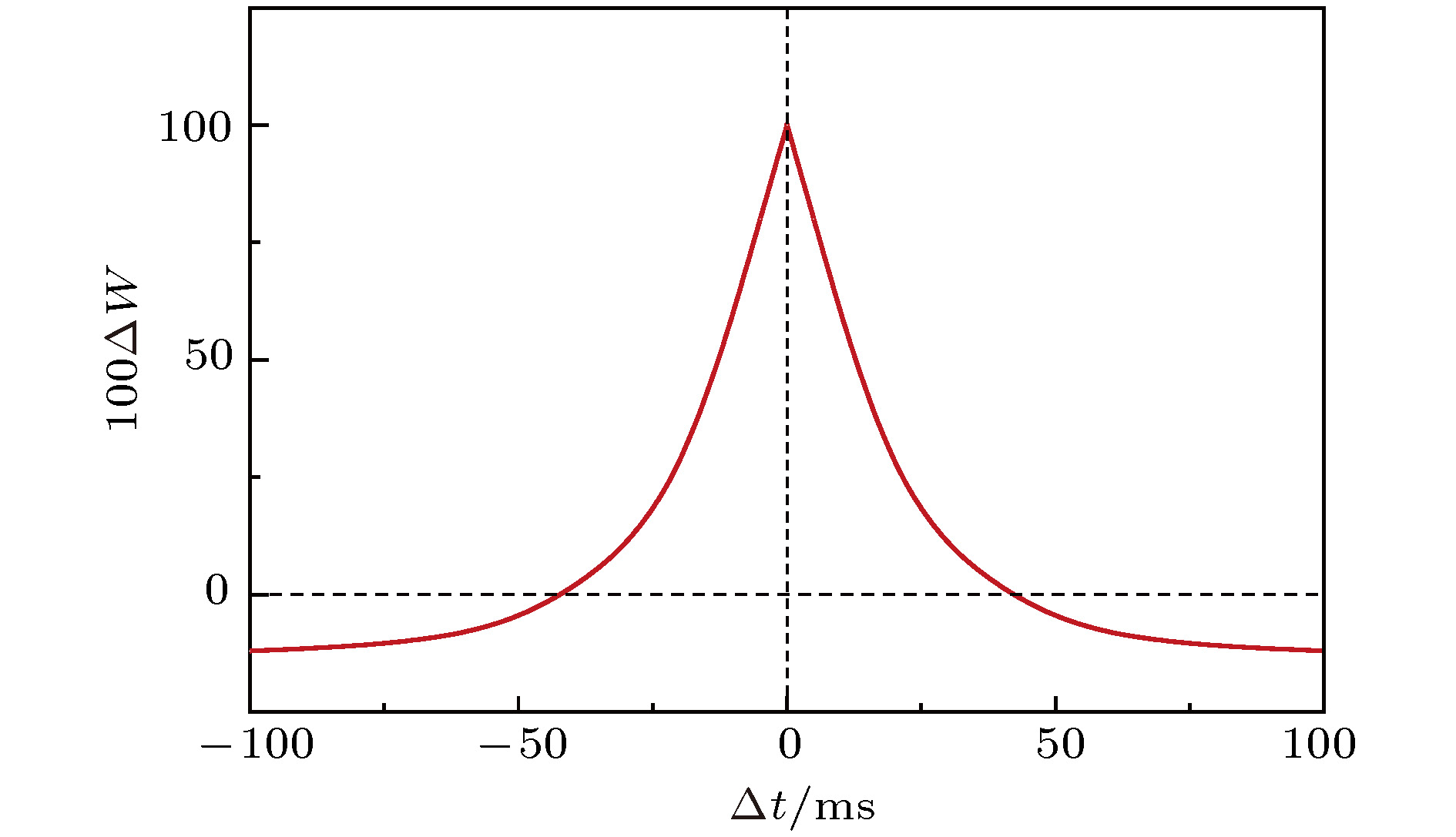
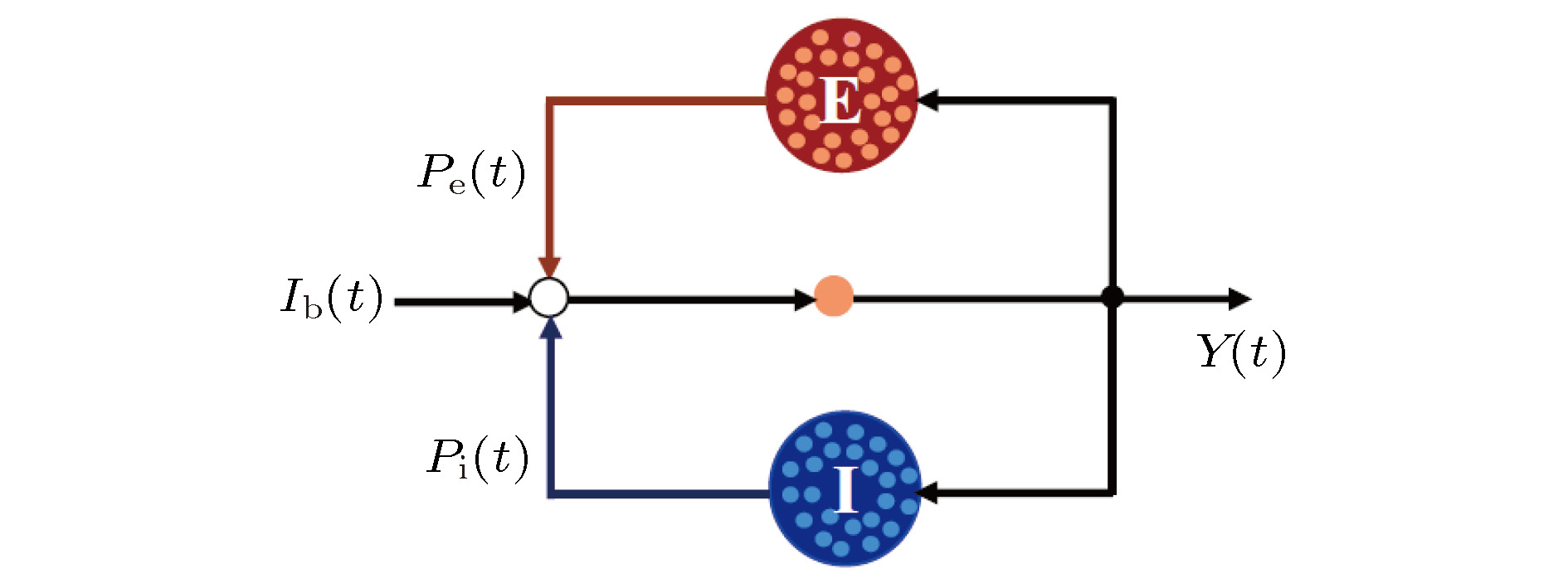
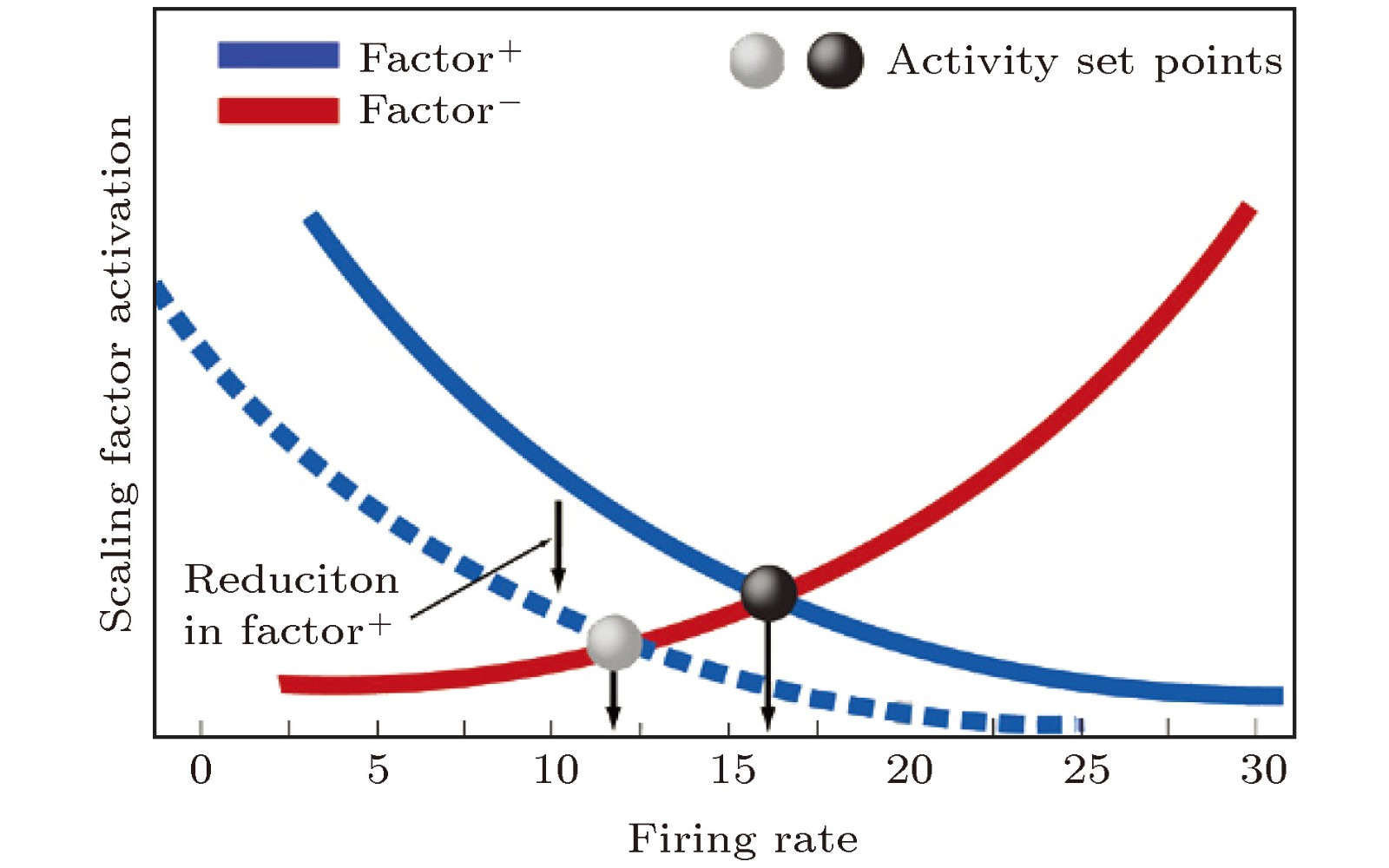
图 2 抑制性突触可塑性(STDP)调节规则示意图
Fig. 2. The regulation rule of inhibitory synaptic plasticity (STDP).
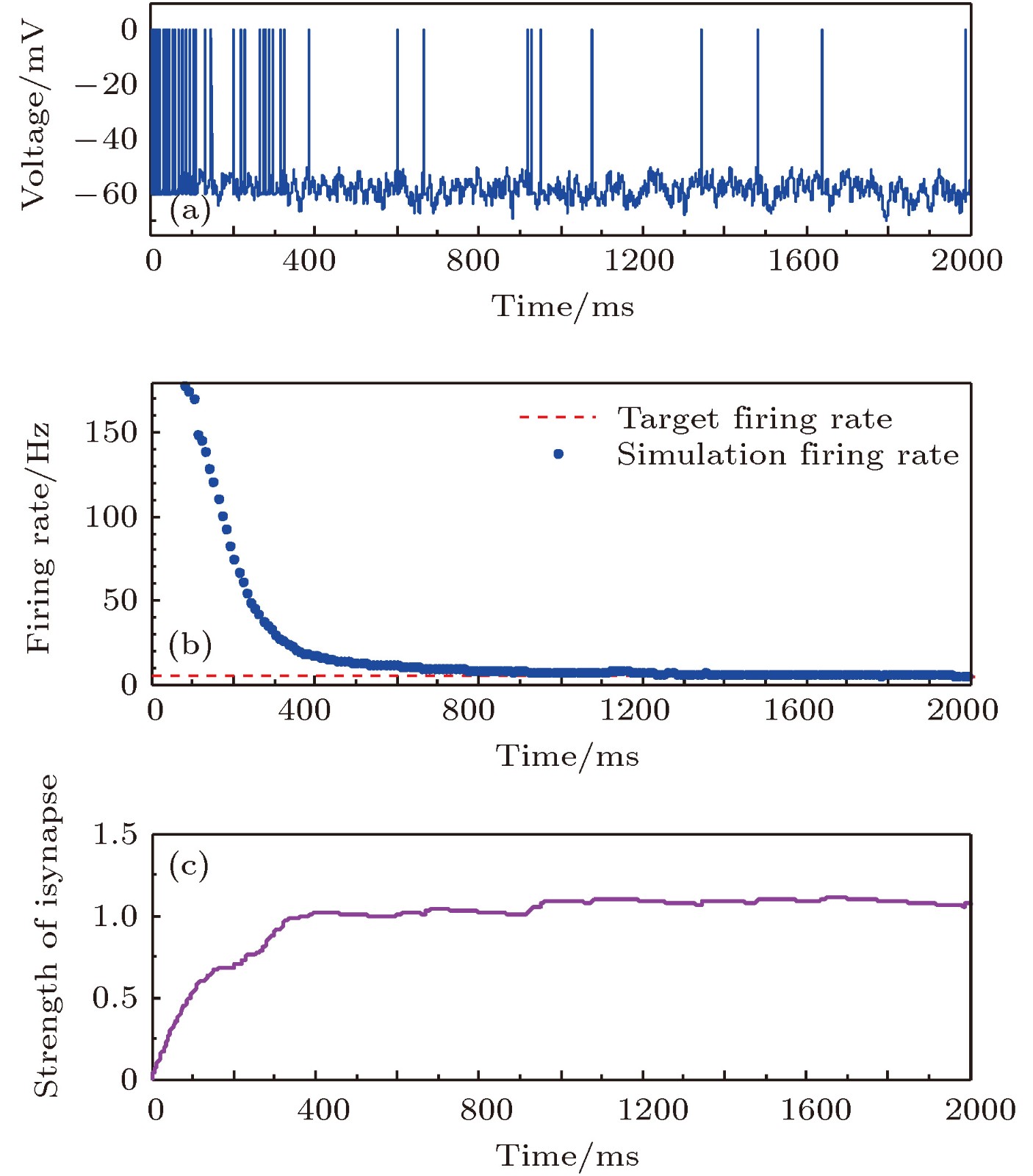
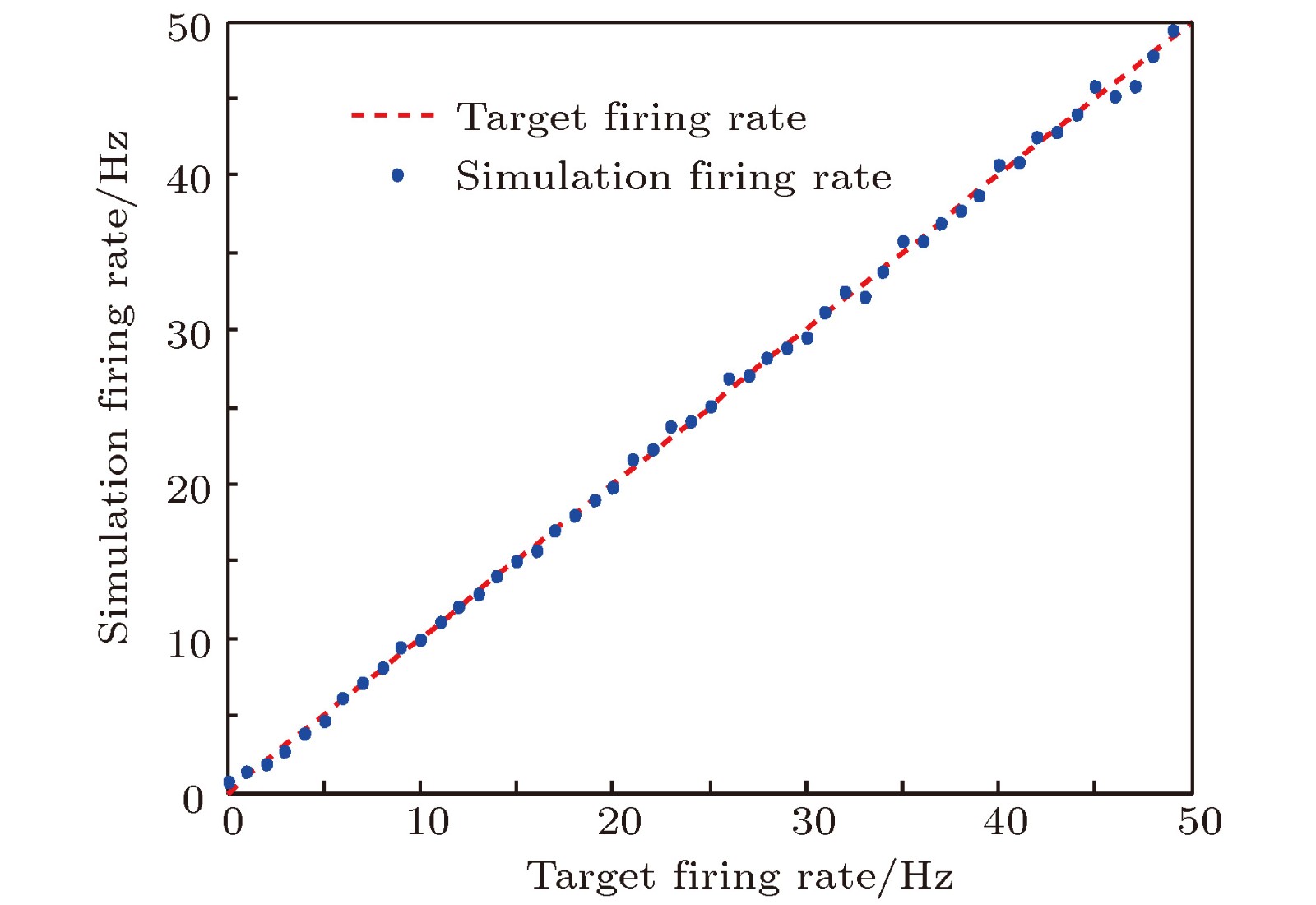
图 3 目标放电率为5 Hz时的神经元放电率自稳态分析图 (a)神经元膜电位; (b)神经元平均放电率曲线; (c)抑制性突触权重变化曲线
Fig. 3. Firing rate homeostasis with the target firing rate equal to 5 Hz: (a) Neural membrane potential; (b) the average firing rate; (c) the strength of inhibitory synapse.
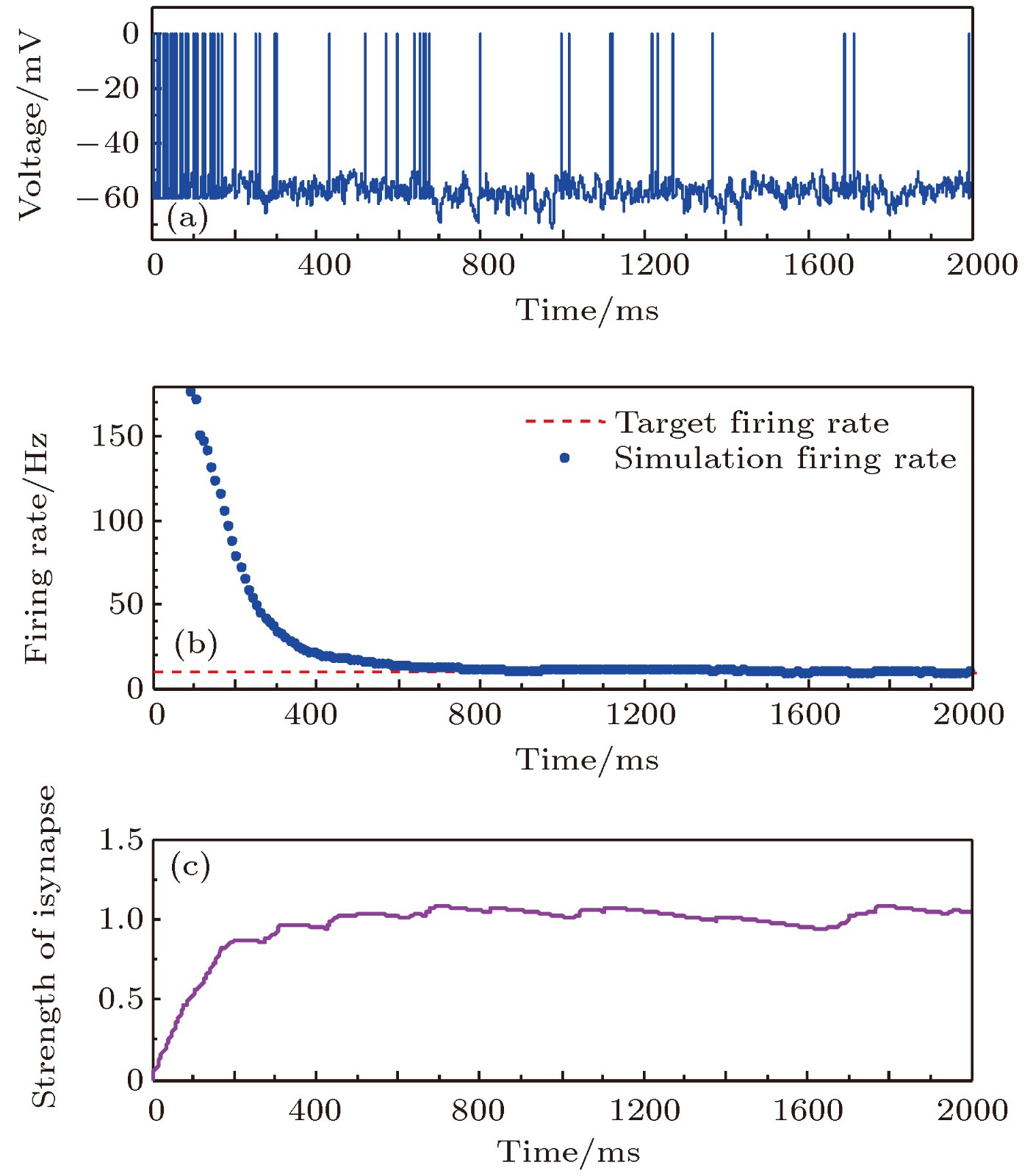
图 4 目标放电率为10 Hz时的神经元放电率自稳态分析图 (a)神经元膜电位; (b)神经元平均放电率曲线; (c)抑制性突触权重变化曲线
Fig. 4. Firing rate homeostasis with the target firing rate equal to 10 Hz: (a) Neuronal membrane potential; (b) the average firing rate; (c) the strength of inhibitory synapse.
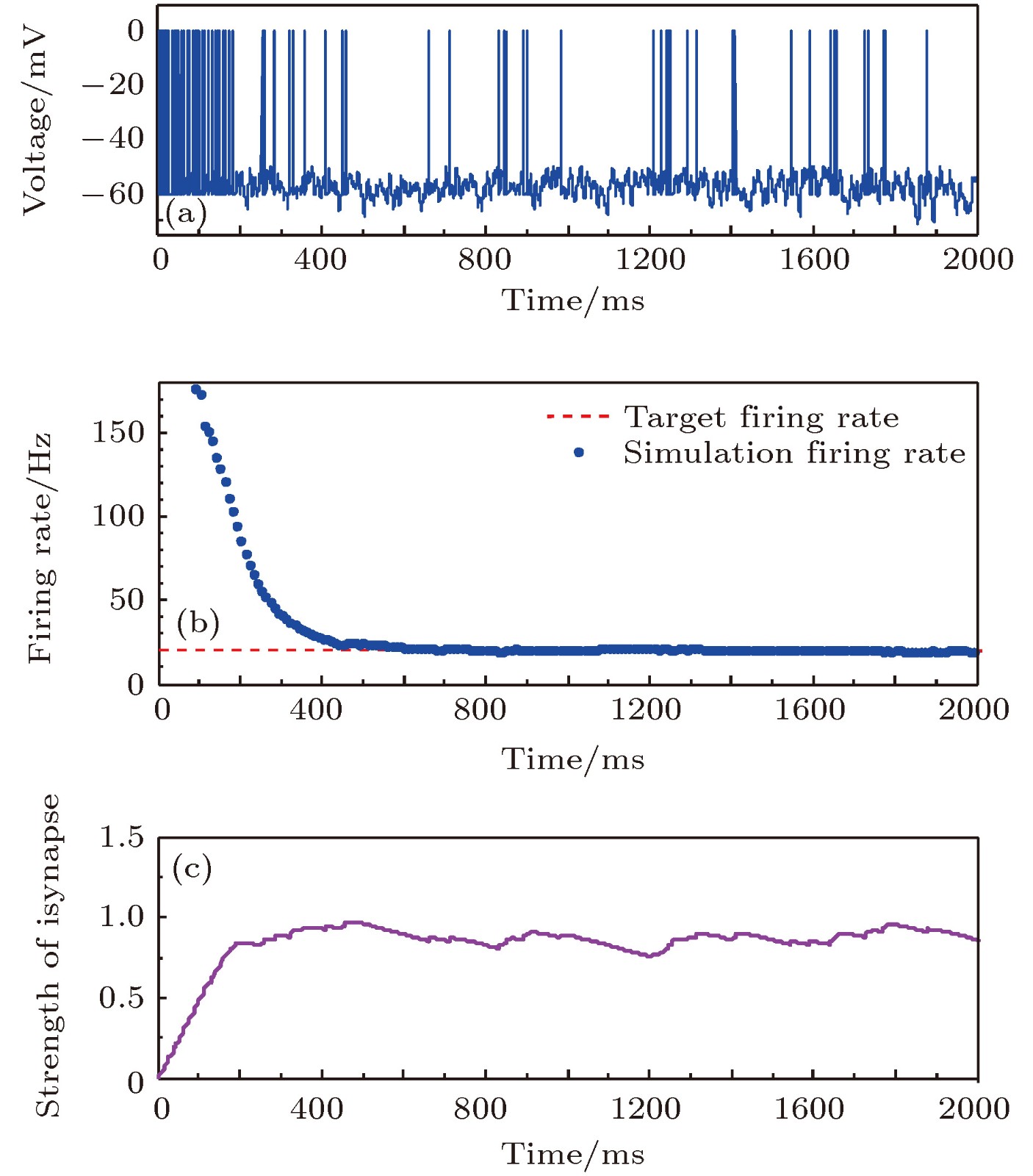
图 5 目标放电率为20 Hz时的神经元放电率自稳态分析图 (a)神经元膜电位; (b)神经元平均放电率曲线; (c)抑制性突触权重变化曲线
Fig. 5. Firing rate homeostasis with the target firing rate equal to 20 Hz: (a) Neuronal membrane potential; (b) the average firing rate; (c) the strength of inhibitory synapse.
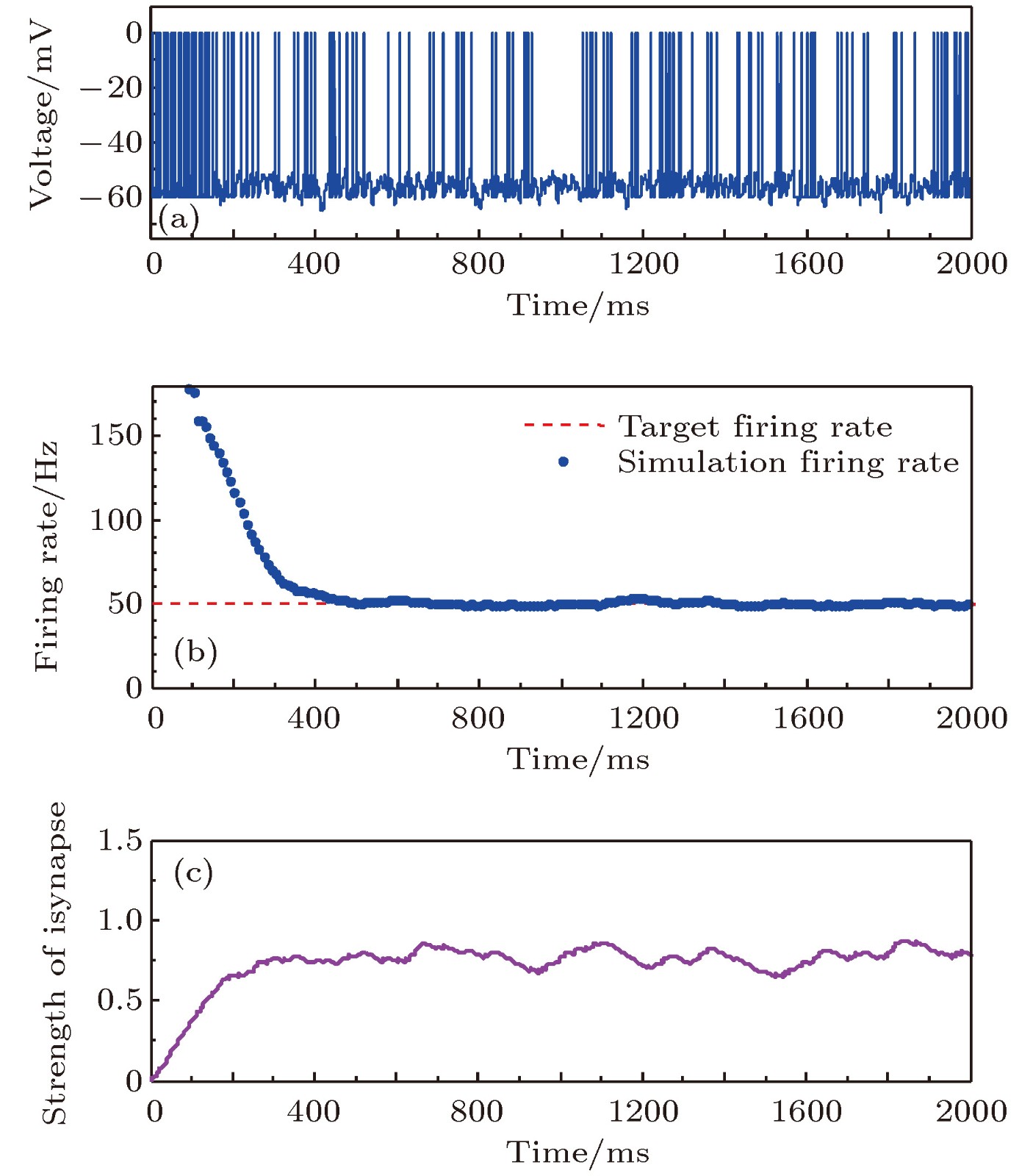
图 6 目标放电率为50 Hz时的神经元放电率自稳态分析图 (a)神经元膜电位; (b)神经元平均放电率曲线; (c)抑制性突触权重变化曲线
Fig. 6. Firing rate homeostasis with the target firing rate equal to 50 Hz: (a) Neuronal membrane potential; (b) the average firing rate; (c) the strength of inhibitory synapse.
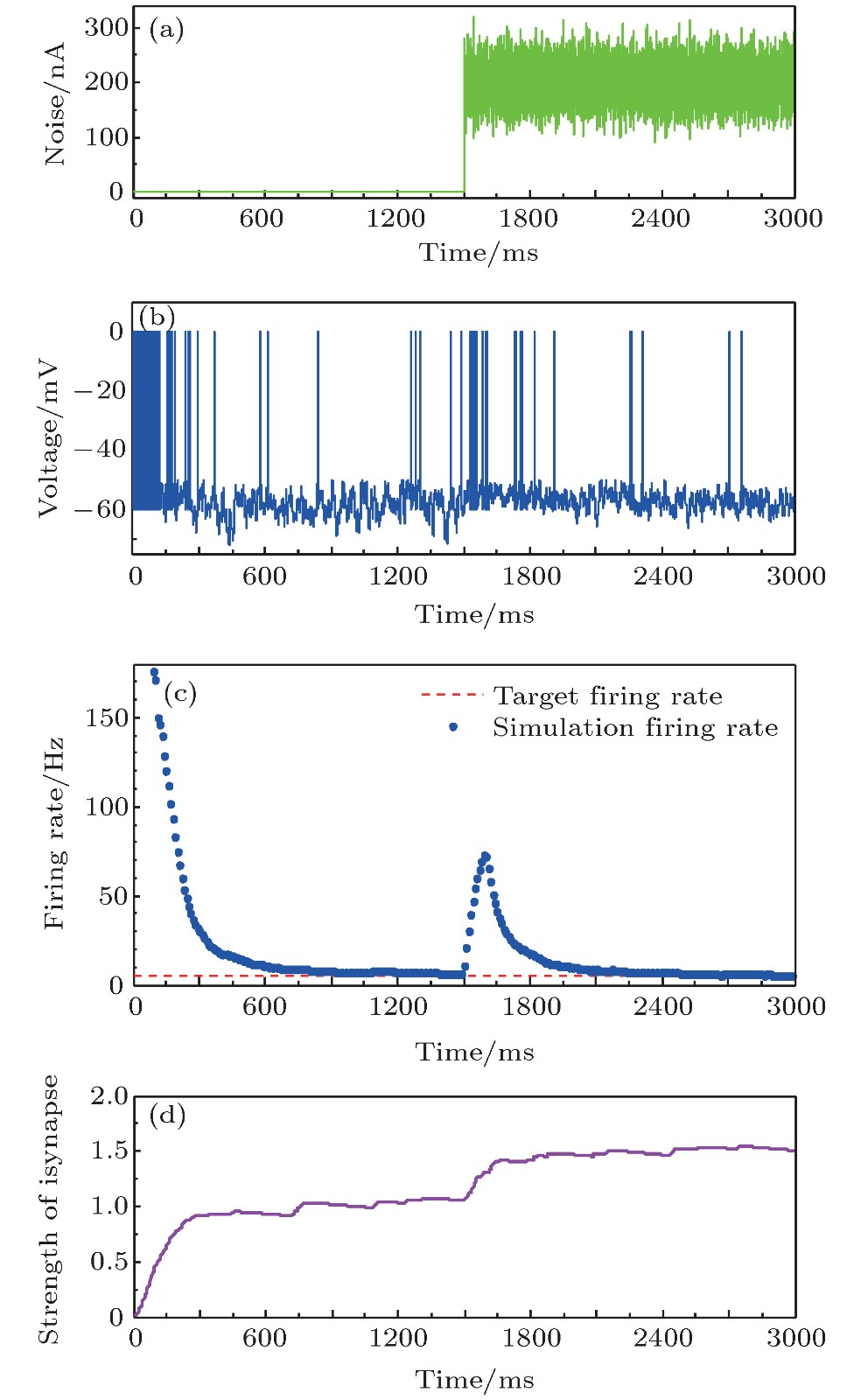
图 8 噪声干扰下的神经元放电率鲁棒性 (a)噪声干扰信号; (b)神经元膜电位; (c)神经元平均放电率曲线; (d)抑制性突触权重变化曲线
Fig. 8. The robustness of neural firing rate homeostasis under noise disturbances: (a) The noise signal; (b) neuronal membrane potential; (c) the average firing rate; (d) the strength of inhibitory synapse.
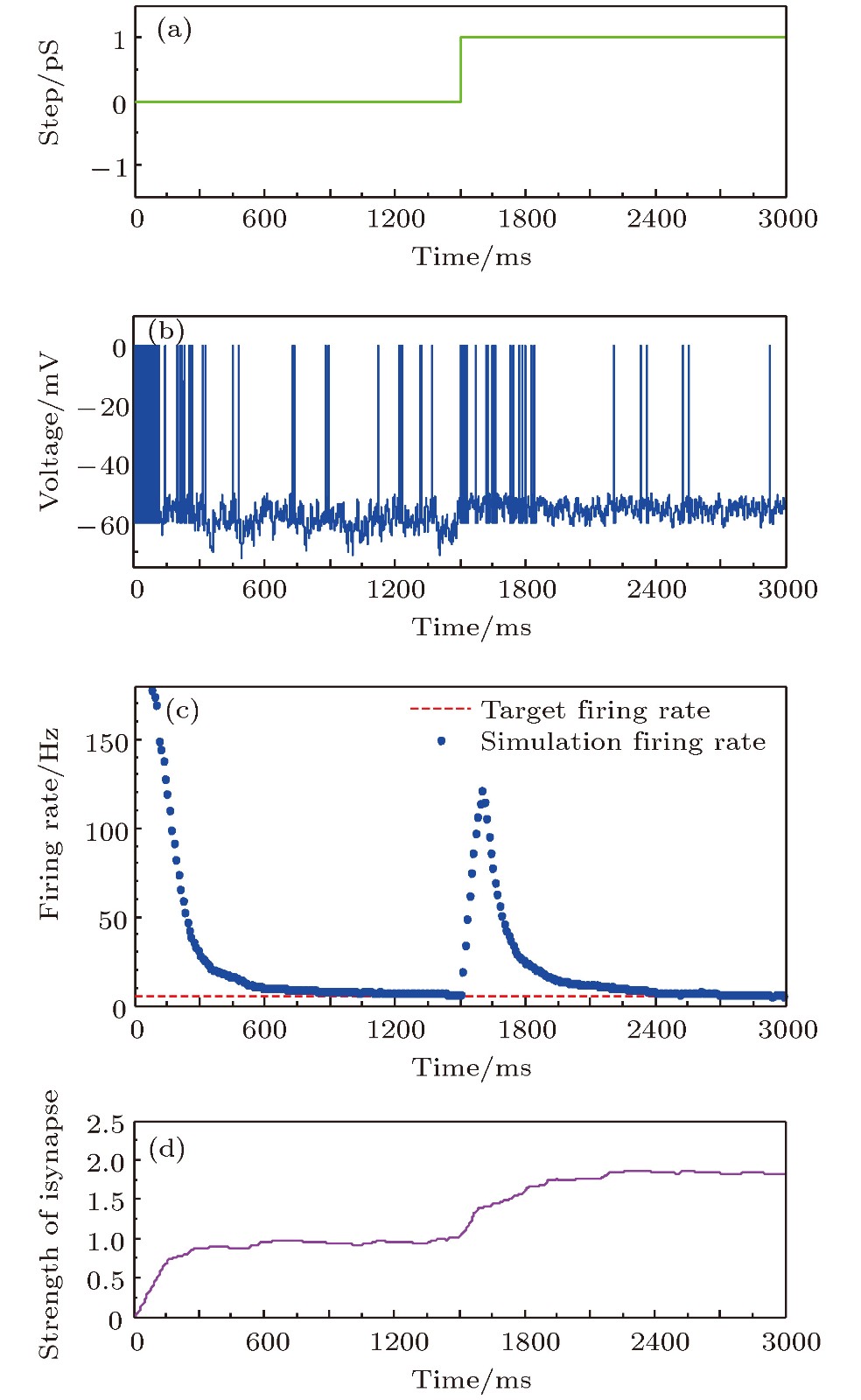
图 9 参数摄动下的放电率自稳态鲁棒性 (a)参数摄动信号; (b)神经元膜电位; (c)神经元平均放电率曲线; (d)抑制性突触权重变化曲线
Fig. 9. The robustness of neural firing rate homeostasis under parameter perturbation: (a) The parameter perturbation signal; (b) neuronal membrane potential; (c) the average firing rate; (d) the strength of inhibitory synapse.
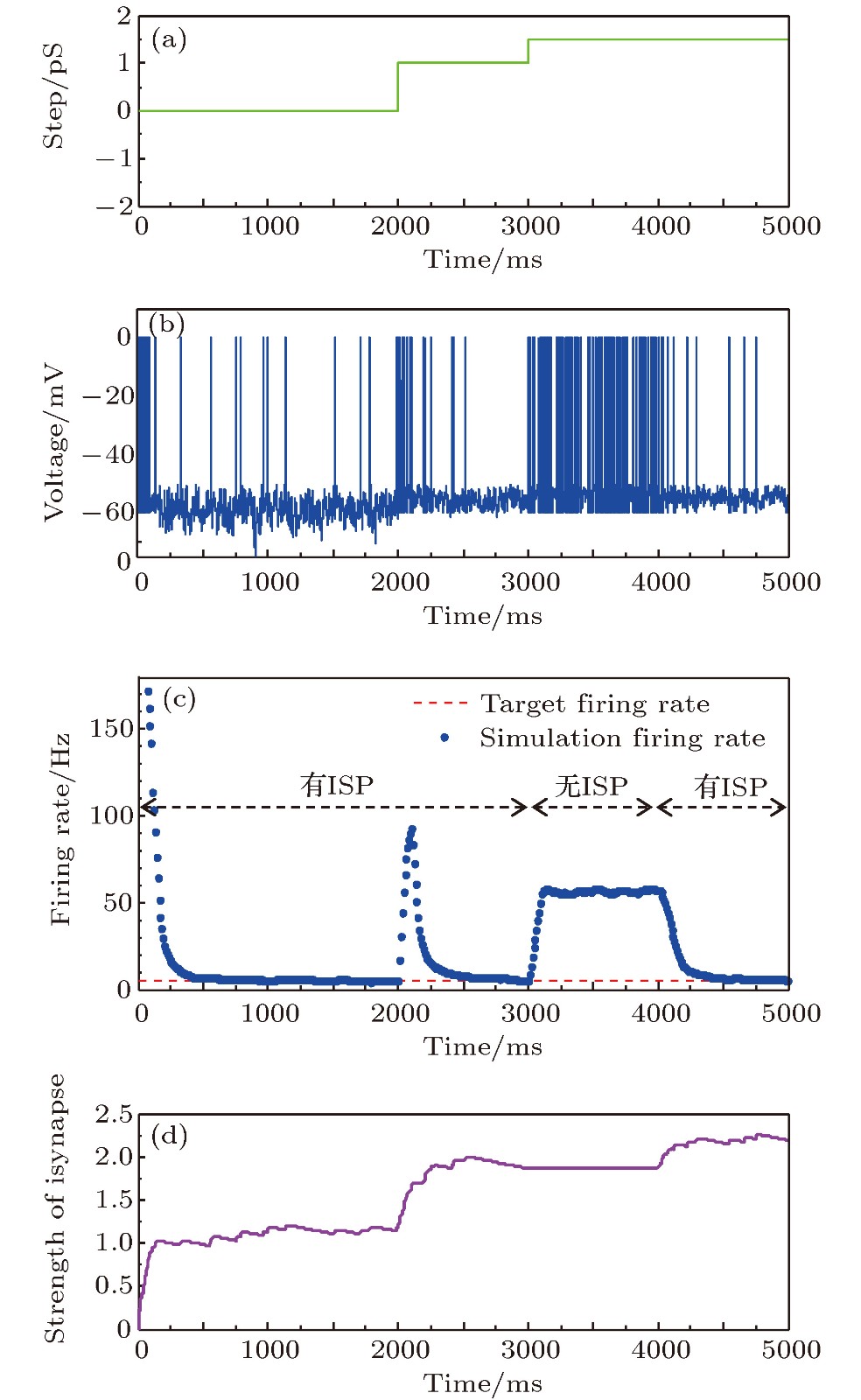
图 10 参数干扰时有无抑制性突触可塑性两种情况下的神经元平均放电率自稳态特性 (a)参数摄动信号; (b)神经元膜电位; (c)神经元平均放电率曲线; (d)抑制性突触权重变化曲线
Fig. 10. The firing rate characteristics with and without inhibitory synaptic plasticity under parameter perturbation: (a) The parameter perturbation signal; (b) neural membrane potential; (c) the average firing rate; (d) the strength of inhibitory synapse.
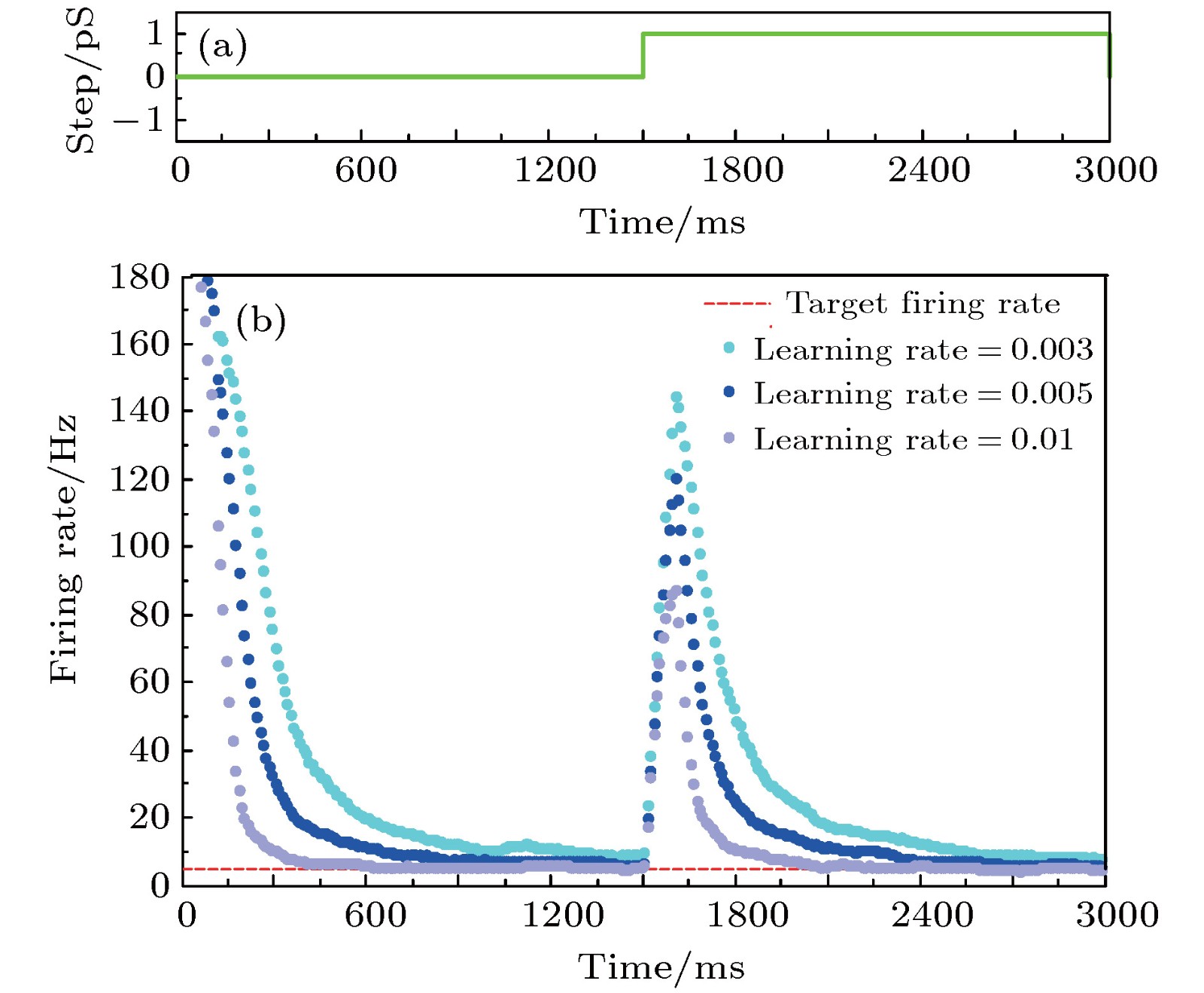
图 11 学习率对放电率自稳态的影响 (a)参数摄动信号; (b)不同学习率时的神经元平均放电率曲线
Fig. 11. The effect of learning rate on neural firing rate homeostasis: (a) The parameter perturbation signal; (b) the firing rate with different learning rates.
表 1 神经网络结构相关参数取值
Table 1. Parameters values of the neural feedback model structure.
参数 描述 取值 NE 兴奋性神经元规模 800 NI 抑制性神经元规模 200 PEE E-E的连接概率 0.2 PEI E-I的连接概率 0.4 PIE I-E的连接概率 0.4 PII I-I的连接概率 0.4  下载: 导出CSV
下载: 导出CSV
表 2 神经元模型各参数取值
Table 2. Parameters values of LIF neuron model.
参数 取值 单位 ${\tau _{\rm{m}}}$ 20 ms ${V^{{\rm{rest}}}}$ –60 mV ${V_{{\rm{th}}}}$ –50 mV ${V^{\rm{E}}}$ 0 mV ${V^{\rm{I}}}$ –70 mV ${g^{{\rm{leak}}}}$ 10 nS
下载: 导出CSV
表 3 突触模型参数取值
Table 3. Parameters values of synapse model.
参数 取值 单位 ${\tau _{\rm{E}}}$ 5 ms ${\tau _{\rm{I}}}$ 10 ms ${\bar g^{\rm{E}}}$ 140 pS ${\bar g^{\rm{I}}}\normalsize$ 350 pS
下载: 导出CSV
表 4 抑制性突触可塑性的参数取值
Table 4. Parameters values of inhibitory synaptic plasticity.
参数 描述 取值 ${W_{ij}}^{}$ 抑制性突触权重 0 ${\tau _{{\rm{STDP}}}}$ 可塑性时间常数 20 $\eta $ 学习率 0.005 $\alpha $ 抑制因子 0.12 ${\rho _0}\normalsize$ 目标放电率
下载: 导出CSV
-
[1] Gläser C, Joublin F 2011 IEEE T. Auton. Ment. De. 3 285
Google Scholar
[2] Hengen K B, Lambo M E, Hooser S D, van Katz D B, Turrigiano G G 2013 Neuron 80 335
Google Scholar
[3] Corner M A, Ramakers G J A 1992 Dev. Brain Res. 65 57
Google Scholar
[4] Ramakers G J A, Corner M A, Habets A M M C 1990 Exp. Brain Res. 79 157
Google Scholar
[5] Ramakers G J A, Galen H V, Feenstra M G P, Corner M A, Boer G J 1994 Int. J. Dev. Neurosci. 12 611
Google Scholar
[6] Pol A N V D, Obrietan K, Belousov A 1996 Neuroscience 74 653
Google Scholar
[7] Turrigiano G G, Leslie K R, Desai N S, Rutherford L C, Nelson S B 1998 Nature 391 892
Google Scholar
[8] Rutherford L C, Nelson S B, Turrigiano G G 1998 Neuron 21 521
Google Scholar
[9] Burrone J, O'Byrne M, Murthy V N 2002 Nature 420 414
Google Scholar
[10] Turrigiano G G, Nelson S B 2004 Nat. Rev. Neurosci. 5 97
Google Scholar
[11] Turrigiano G 2012 CSH Perspect. Biol. 4 a005736
Google Scholar
[12] Cannon J, Miller P 2016 J. Neurophysiol. 116 2004
Google Scholar
[13] Cannon J, Miller P 2017 J. Math. Neurosc. 7 1
Google Scholar
[14] Miller P, Cannon J 2018 Biol. Cybern. 113 47
[15] McClelland J L, McNaughton B L, O'Reilly R C 1995 Psychol. Rev. 102 419
Google Scholar
[16] Frankland P W, O'Brien C, Ohno M, Kirkwood A, Silva A J 2001 Nature 411 309
Google Scholar
[17] Carcea I, Froemke R C 2013 Prog. Brain. Res. 207 65
Google Scholar
[18] Martin S J, Grimwood P D, Morris R G M 2000 Annu. Rev. Neurosci. 23 649
Google Scholar
[19] Sanderson J L, Dell'Acqua M L 2011 Neuroscientist 17 321
Google Scholar
[20] Yong L, Kauer J A 2010 Synapse 51 1
Google Scholar
[21] Haas J S, Thomas N, Abarbanel H D I 2006 J. Neurophysiol. 96 3305
Google Scholar
[22] D'Amour J A, Froemke R C 2015 Neuron 86 514
Google Scholar
[23] Hartmann K, Bruehl C, Golovko T, Draguhn A 2008 Plos One 3 e2979
Google Scholar
[24] Tohru K, Kazumasa Y, Yumiko Y, Crair M C, Yukio K 2008 Neuron 57 905
Google Scholar
[25] Stephen G, James R W 2001 Cereb. Cortex 11 37
Google Scholar
[26] Luz Y, Shamir M 2012 Plos. Comput. Biol. 8 e1002334
Google Scholar
[27] Hennequin G, Agnes E J, Vogels T P 2017 Annu. Revi. Neurosci. 40 557
Google Scholar
[28] Park H J, Friston K 2013 Science 342 1238411
Google Scholar
[29] Isaacson J S, Massimo S 2011 Neuronv 72 231
Google Scholar
[30] Maass W, Joshi P, Sontag E D 2007 Plos Comput. Biol. 3 e165
Google Scholar
[31] Jansen B H, Rit V G 1995 Biol. Cybern. 73 357
Google Scholar
[32] Chacron M J, André L, Leonard M 2005 Phys. Rev. E 72 051917
Google Scholar
[33] Froemke R C, Jones B J 2011 Neurosci. Biobehav. R. 35 2105
Google Scholar
[34] 王俊松, 徐瑶 2014 物理学报 63 068701
Google Scholar
Wang J S, Xu Y 2014 Acta Phys. Sin. 63 068701
Google Scholar
[35] 王美丽, 王俊松 2015 物理学报 64 108701
Google Scholar
Wang M L, Wang J S 2015 Acta Phys. Sin. 64 108701
Google Scholar
[36] Vogels T P, Abbott L F 2009 Nat. Neurosci. 12 483
Google Scholar
[37] Stepp N, Plenz D, Srinivasa N 2015 Plos Comput. Biol. 11 e1004043
Google Scholar
[38] Vogels T P, Sprekeler H, Zenke F, Clopath C, Gerstner W 2011 Science 334 1569
Google Scholar
[39] Maass W 2014 P. IEEE 102 860
Google Scholar
[40] Mcdonnell M D, Ward L M 2011 Nat. Rev. Neurosci. 12 183
Google Scholar
[41] Garrett D D, Mcintosh A R, Grady C L 2011 Nat. Rev. Neurosci. 12 612
Google Scholar
[42] Mcdonnell M D, Ward L M 2011 Nat. Rev. Neurosci. 12 415
Google Scholar
[43] Turrigiano G G 2008 Cell 135 422
Google Scholar
[44] Marder E, Tang L S 2010 Neuron 66 161
Google Scholar
[45] Sharon B, Dickman D K, Davis G W 2010 Neuron 66 220
Google Scholar

 下载:
下载:

计量
- 文章访问数: 24097
- PDF下载量: 148
- 被引次数: 0
