










-
Pre-Bötzinger复合体是兴奋性耦合的神经元网络, 通过产生复杂的放电节律和节律模式的同步转迁参与调控呼吸节律. 本文选用复杂簇和峰放电节律的单神经元数学模型构建复合体模型, 仿真了与生物学实验相关的多类同步节律模式及其复杂转迁历程, 并利用快慢变量分离揭示了相应的分岔机制. 当初值相同时, 随着兴奋性耦合强度的增加, 复合体模型依次表现出完全同步的“fold/homoclinic”, “subHopf/subHopf”簇放电和周期1峰放电. 当初值不同时, 随耦合强度增加, 表现为由“fold/homoclinic”, 到“fold/fold limit cycle”、到“subHopf/subHopf”与“fold/fold limit cycle”的混合簇放电、再到“subHopf/subHopf”簇放电的相位同步转迁, 最后到反相同步周期1峰放电. 完全(同相)同步和反相同步的周期1节律表现出了不同分岔机制. 反相峰同步行为给出了与强兴奋性耦合容易诱发同相同步这一传统观念不同的新示例. 研究结果给出了pre-Bötzinger复合体的从簇到峰放电节律的同步转迁规律及复杂分岔机制, 反常同步行为丰富了非线性动力学的内涵.
-
关键词:
- 分岔 /
- 同步转迁 /
- 神经放电 /
- Pre-Bötzinger复合体
The pre-Bötzinger complex is a neuronal network with excitatory coupling, which participates in modulation of respiratory rhythms via the generation of complex firing rhythm patterns and synchronization transitions of rhythm patterns. In the present paper, a mathematical model of single neuron that exhibits complex transition processes from bursting to spiking is selected as a unit, the network model of the pre-Bötzinger complex composed of two neurons with excitatory coupling is constructed, multiple synchronous rhythm patterns and complex transition processes of the synchronous rhythm patterns related to the biological experimental observations are simulated, and the corresponding bifurcation mechanism is acquired with the fast-slow variable dissection method. When the initial values of two neurons of the pre-Bötzinger complex are the same, with increasing the excitatory coupling strength, the theoretical model of the pre-Bötzinger complex shows complete synchronization transition processes from "fold/homoclinic" bursting, to "subHopf/subHopf" bursting, and at last to period-1 spiking. When the initial values are different, with the increases of the excitatory coupling intensity, the rhythm transition processes begin from phase synchronization behaviors including "fold/homoclinic" bursting, "fold/fold limit cycle" bursting, mixed bursting composed of "subHopf/subHopf" bursting and "fold/fold limit cycle" bursting, and "subHopf/ subHopf" bursting in sequence, and to anti-phase synchronous behavior of the period-1 spiking. The complete (in-phase) synchronous period-1 spiking for the same initial values exhibits bifurcation mechanism different from the anti-phase synchronous period-1 spiking for different initial values. The anti-phase synchronous period-1 spiking presents a novel and abnormal example of the synchronization at large excitatory coupling strength, which is different from the traditional viewpoint that large excitatory coupling often induces in-phase synchronous behavior. The results present the synchronization transition process and complex bifurcation mechanism from bursting to period-1 spiking of the pre-Bötzinger complex, and the abnormal synchronization example enriches the contents of nonlinear dynamics.-
Keywords:
- bifurcation /
- synchronization transition /
- neural firing /
- pre-Bötzinger complex
[1] Braun H A, Wissing H, Schäfer K, Hirsch M C 1994 Nature 367 270
 Google Scholar
Google Scholar
[2] Gu H G, Pan B B 2015 Nonlinear Dyn. 81 2107
Google Scholar
[3] 谢勇, 程建慧 2017 物理学报 66 090501
Google Scholar
Xie Y, Cheng J H 2017 Acta Phys. Sin. 66 090501
Google Scholar
[4] Sun X J, Perc M, Kurths J, Lu Q S 2018 Chaos 28 106310
Google Scholar
[5] 徐莹, 王春妮, 靳伍银, 马军 2015 物理学报 64 198701
Xu Y, Wang C N, Jin W Y, Ma J 2015 Acta Phys. Sin. 64 198701
[6] 李国芳, 孙晓娟 2017 物理学报 66 240501
Google Scholar
Li G F, Sun X J 2017 Acta Phys. Sin. 66 240501
Google Scholar
[7] Bianchi A L, Denavit-Saubie M, Champagnat J 1995 Physiol. Rev. 75 1
Google Scholar
[8] Cohen M I 1979 Physiol. Rev. 59 1105
Google Scholar
[9] Funk G D, Smith J C, Feldman J L 1995 J. Neurosci. 15 4046
Google Scholar
[10] Richter D W, Ballanyi K, Schwarzacher S 1992 Curr. Opin. Neurobiol. 2 788
Google Scholar
[11] 严亨秀, 张承武, 郑煜 2004 生理学报 56 665
Google Scholar
Yan H X, Zhang C W, Zheng Y 2004 Acta Physiol. Sin. 56 665
Google Scholar
[12] 宋刚 1999 生理科学进展 3 237
Song G 1999 Prog. Physiol. Sci. 3 237
[13] Smith J C, Ellenberger H H, Ballanyi K, Richter D W, Feldman J L 1991 Science 254 726
Google Scholar
[14] Feldman J L, Negro C A D 2006 Nat. Rev. Neurosci. 7 232
Google Scholar
[15] Smith J C 1997 Neurons, Networks, and Motor Behavior (Cambridge, MA: MIT Press) p97
[16] Johnson S M, Smith J C, Funk G D, Feldman J L 1994 J. Neurophysiol. 72 2598
Google Scholar
[17] Ramirez J M, Richter D W 1996 Curr. Opin. Neurobiol. 6 817
Google Scholar
[18] Rekling J C, Feldman J L 1998 Annu. Rev. Physiol. 60 385
Google Scholar
[19] Koshiya N, Smith J C 1998 28th Annual Meeting of the Society for Neuroscience Los Angeles, California, USA, November 7-12, 1998 p531
[20] Koshiya N, Smith J C 1999 Nature 400 360
Google Scholar
[21] Negro C A D, Morgado V C, Hayes J A, Mackay D D, Pace R W, Crowder E A, Feldman J L 2005 J. Neurosci. 25 446
Google Scholar
[22] Smith J C, Butera R J, Koshiya N, Del Negro C, Wilson C G, Johnson S M 2000 Resp. Physiol. 122 131
Google Scholar
[23] Gray P A, Rekling J C, Bocchiaro C M, Feldman J L 1999 Science 286 1566
Google Scholar
[24] Butera R J, Rinzel J, Smith J C 1999 J. Neurophysiol. 82 382
Google Scholar
[25] Dunmyre J R, Negro C A D, Rubin J E 2011 J. Comput. Neurosci. 31 305
Google Scholar
[26] Negro C A D, Johnson S M, Butera R J, Smith J C 2001 J. Neurophysiol. 86 59
Google Scholar
[27] Butera R J, Rinzel J, Smith J C 1999 J. Neurophysiol. 82 398
Google Scholar
[28] Purvis L K, Smith J C, Koizumi H, Butera R J 2007 J. Neurophysiol. 97 1515
Google Scholar
[29] Best J, Borisyuk A, Rubin J E, Terman D, Wechselberger M 2005 SIAM J. Appl. Dyn. Syst. 4 1107
Google Scholar
[30] Rubin J E 2006 Phys. Rev. E 74 021917
Google Scholar
[31] Dunmyre J R, Rubin J E 2010 SIAM J Appl. Dyn. Syst. 9 154
Google Scholar
[32] Guo D D, Lü Z S 2019 Chin. Phys. B 28 110501
Google Scholar
[33] Rybak I A, Molkov Y I, Jasinski P E, Shevtsova N A, Smith J C 2014 Prog. Brain. Res. 209 1
Google Scholar
[34] 张应腾, 熊冬生, 刘深泉 2015 中国医学物理学杂志 32 115
Google Scholar
Zhang Y T, Xiong D S, Liu S Q 2015 Chin. J. Med. Phys. 32 115
Google Scholar
[35] 刘义, 刘深泉 2011 动力学与控制学报 9 257
Google Scholar
Liu Y, Liu S Q 2011 J. Dynam. Cont. 9 257
Google Scholar
[36] Duan L X, Zhai D H, Tang X H 2012 Int. J. Bifurcation Chaos 22 1250114
Google Scholar
[37] Lü Z S, Chen L N, Duan L X 2019 Appl. Math. Model. 67 234
Google Scholar
[38] Lü Z S, Zhang B Z, Duan L X 2017 Cogn. Neurodynamics 11 443
Google Scholar
[39] Wang Z J, Duan L X, Cao Q Y 2018 Chin. Phys. B 27 070502
Google Scholar
[40] Duan L X, Liu J, Chen X, Xiao P C, Zhao Y 2017 Cogn. Neurodynamics 11 91
Google Scholar
[41] Rubin J E, Shevtsova N A, Ermentrout G B, Smith J C, Rybak I A 2009 J. Neurophysiol. 101 2146
Google Scholar
[42] Rubin J E, Bacak B J, Molkov Y I, Shevtsova N A, Smith J C, Rybak I A 2011 J. Comput. Neurosci. 30 607
Google Scholar
[43] 平小方, 刘深泉, 任会霞 2015 动力学与控制学报 13 215
Google Scholar
Ping X F, Liu S Q, Ren H X 2015 J. Dynam. Cont. 13 215
Google Scholar
[44] Belykh I, Shilnikov A 2008 Phys. Rev. Lett. 101 078102
Google Scholar
[45] Wu F Q, Gu H G, Li Y Y 2019 Commun. Nonlinear Sci. Numer. Simul. 79 104924
Google Scholar
[46] 丁学利, 李玉叶 2016 物理学报 65 210502
Google Scholar
Ding X L, Li Y Y 2016 Acta Phys. Sin. 65 210502
Google Scholar
[47] Jia B, Wu Y C, He D, Guo B H, Xue L 2018 Nonlinear Dyn. 93 1599
Google Scholar
[48] Li Y Y, Gu H G, Ding X L 2019 Nonlinear Dyn. 97 2091
Google Scholar
[49] 曹奔, 关利南, 古华光 2018 物理学报 67 240502
Google Scholar
Cao B, Guan L N, Gu H G 2018 Acta Phys. Sin. 67 240502
Google Scholar
[50] Uzuntarla M, Torres J J, Calim A, Barreto E 2019 Neural Networks 110 131
Google Scholar
[51] 埃门创特 B 著 (孝鹏程, 段丽霞, 苏建忠译) 2002 动力系统仿真, 分析与动画—XPPAUT使用指南 (北京: 科学出版社) 第155−167页
Ermentrout B (translated by Xiao P C, Duan L L, Su J Z) 2002 Simulating, Analyzing, and Animating Dynamical systems: A Guide to XPPAUT for Researchers and Students (Beijing: Science Press) p155−167 (in Chinese)
[52] Izhikevich E M 2000 Int. J. Bifurcation Chaos 10 1171
Google Scholar
[53] Hashemi M, Valizadeh A, Azizi Y 2012 Phys. Rev. E 85 021917
Google Scholar
[54] Jia B, Gu H G 2017 Int. J. Bifurcation Chaos 27 1750113
Google Scholar
[55] Zhao Z G, Gu H G 2017 Sci. Rep. 7 6760
Google Scholar
[56] Li J J, Du M M, Wang R, Lei J Z, Wu Y 2016 Int. J. Bifurcation Chaos 26 1650138
Google Scholar
-

图 1 不同
${g_{\rm{K}}}$ 下单神经元放电在(h, V)相平面的轨迹 (a)$ {g_{\rm{K}}} = {\rm{7}}{\rm{.1}}\;{\rm{nS}} $ ; (b)$ {g_{\rm{K}}} = {\rm{7}}{\rm{.8}}\;{\rm{nS}} $ ; (c)$ {g_{\rm{K}}} = {\rm{10}}{\rm{.0}}\;{\rm{nS}} $ ; (d)$ {g_{\rm{K}}} = {\rm{25}}{\rm{.0}}\;{\rm{nS}} $ Fig. 1. The (h, V) trajectory of the single neuron at different
${g_{\rm{K}}}$ values: (a)$ {g_{\rm{K}}} = {\rm{7}}{\rm{.1}}\;{\rm{nS}} $ ; (b)$ {g_{\rm{K}}} = {\rm{7}}{\rm{.8}}\;{\rm{nS}} $ ; (c)$ {g_{\rm{K}}} = {\rm{10}}{\rm{.0}}\;{\rm{nS}} $ ; (d)$ {g_{\rm{K}}} = {\rm{25}}{\rm{.0}}\;{\rm{nS}} $ .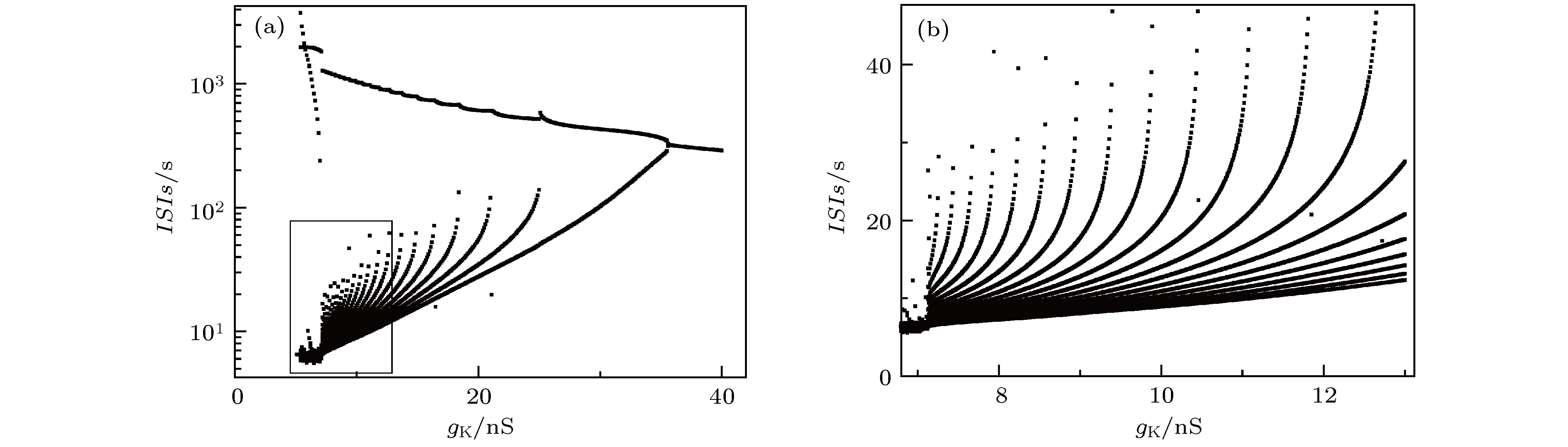
图 2 单神经元模型的随
${g_{\rm{K}}}$ 的分岔 (a) ISIs分岔序列; (b)图(a)左下角方框的局部放大Fig. 2. Bifurcation of the single neuron model with increasing
${g_{\rm{K}}}$ : (a) Bifurcations of ISIs; (b) the enlargement of ISIs within the square at the down-left corner of fig (a).
图 3 当
$ {g_{\rm{K}}} = 7.1\;{\rm{nS}} $ 时, 单神经元模型的快子系统随着慢变量h变化的分岔Fig. 3. Bifurcations of the fast-subsystem of the single neuron with respect to h when
$ {g_{\rm{K}}} = 7.1\;{\rm{nS}} $ .
图 4 单神经元在不同的
${g_{\rm{K}}}$ 下簇放电模式的快慢变量分离 (a)$ {g_{\rm{K}}} = {\rm{7}}.1\;{\rm{nS}} $ ; (b)$ {g_{\rm{K}}} = {\rm{7}}.8\;{\rm{nS}} $ ; (c)$ {g_{\rm{K}}} = {\rm{10}}.0\;{\rm{nS}} $ ; (d)${g_{\rm{K}}} =$ 25.0 nSFig. 4. The fast-slow variable dissection of bursting of single neuron at different
${g_{\rm{K}}}$ values: (a)$ {g_{\rm{K}}} = {\rm{7}}.1\;{\rm{nS}} $ ; (b)$ {g_{\rm{K}}} = {\rm{7}}.8\;{\rm{nS}} $ ; (c)$ {g_{\rm{K}}} = {\rm{10}}.0\;{\rm{nS}} $ ; (d)$ {g_{\rm{K}}} = {\rm{25}}.0\;{\rm{nS}} $ .
图 5 随着耦合强度
${g_{{\text{syn-e}}}}$ 增大, 耦合神经元模型的同步转迁过程. 相同初值 (a1)耦合电流平均值$\bar I$ ; (a2)峰相位差的最大值$\max (\Delta (\phi (t)))$ ; (a3)簇相位差的最大值$ \max (\Delta (\varPhi (t))) $ ; (a4)相关系数ρ; (a5)神经元1的ISIs序列. 不同初值: (b1)耦合电流平均值$\bar I$ ; (b2)峰相位差的最大值$\max (\Delta (\phi (t)))$ ; (b3)簇相位差的最大值$ \max (\Delta (\varPhi (t))) $ ; (b4)相关系数ρ; (b5)神经元1的ISIs序列Fig. 5. Transitions with respect to
${g_{{\text{syn-e}}}}$ of coupled neurons model. The same initial values: (a1) The mean values of coupling current$\bar I$ ; (a2) maximum spike phase difference$\max (\Delta (\phi (t)))$ ; (a3) maximum burst phase difference$ \max (\Delta (\varPhi (t))) $ ; (a4) coefficient ρ; (a5) ISIs of neuron 1. Different initial values: (b1) The mean values of coupling current$\bar I$ ; (b2) maximum spike phase difference$\max (\Delta (\phi (t)))$ ; (b3) maximum burst phase difference$ \max (\Delta (\varPhi (t))) $ ; (b4) coefficient ρ; (b5) ISIs of neuron 1.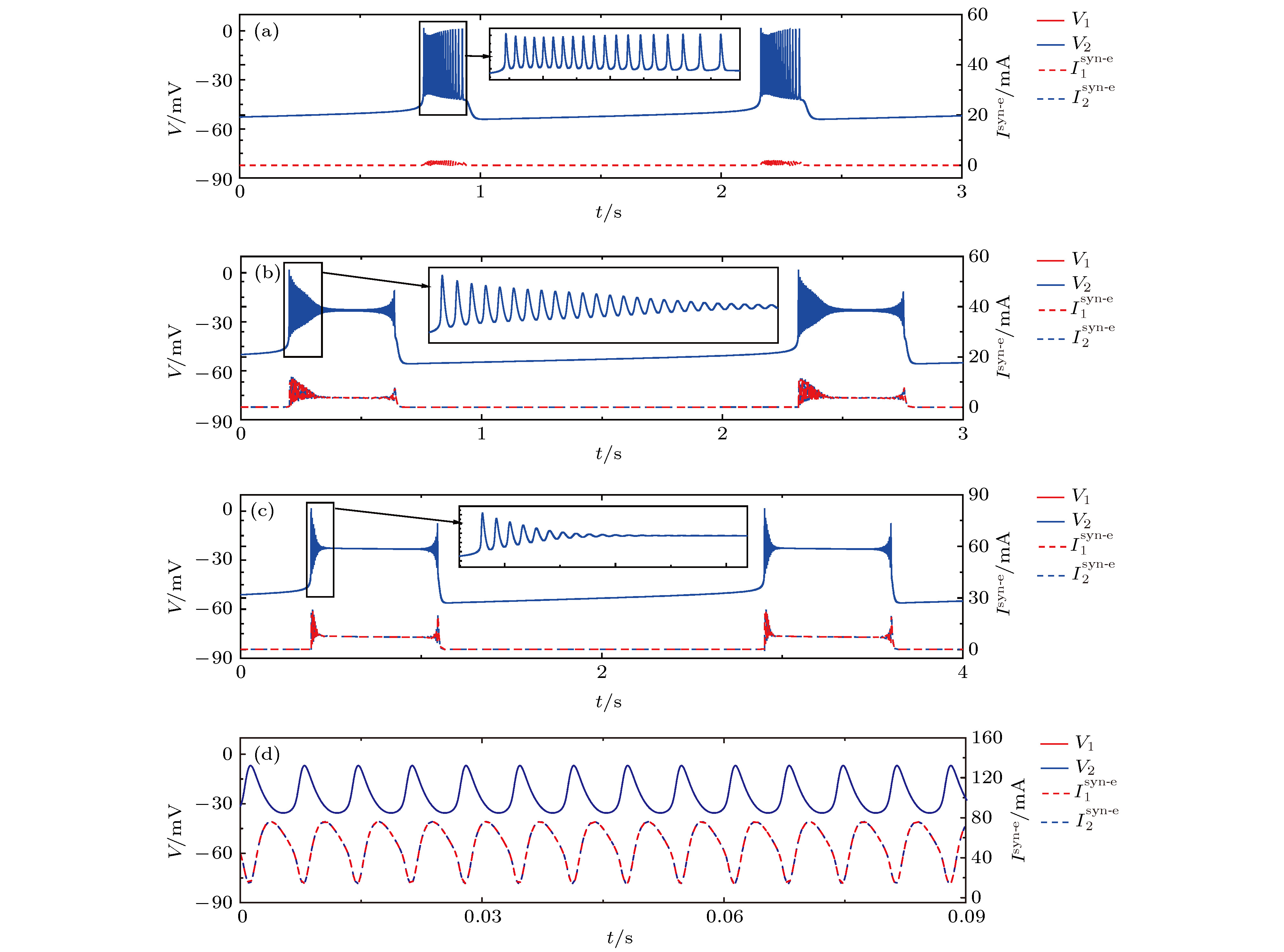
图 6 初值相同时, 不同耦合强度下神经元1(红)和2(蓝)的膜电位
$V$ (上)及耦合电流${I^{{\text{syn-e}}}}$ (下), 插图是局部放大 (a)$ {g_{{\text{syn-e}}}} = {\rm{0}}{\rm{.35}}\;{\rm{nS}} $ ; (b)$ {g_{{\text{syn-e}}}} = {\rm{2}}{\rm{.5}}\;{\rm{nS}} $ ; (c)$ {g_{{\text{syn-e}}}} = {\rm{5}}{\rm{.0}}\;{\rm{nS}} $ ; (d)$ {g_{{\text{syn-e}}}} = {\rm{18}}{\rm{.0}}\;{\rm{nS}} $ Fig. 6. Membrane potential
$V$ (top) and coupling current${I^{{\text{syn-e}}}}$ (low) of neurons 1 (red) and 2 (blue) with the same initial values at different${g_{{\text{syn-e}}}}$ values (Insert figure: the enlargement of bursting): (a)$ {g_{{\text{syn-e}}}} = {\rm{0}}{\rm{.35}}\;{\rm{nS}} $ ; (b)$ {g_{{\text{syn-e}}}} = {\rm{2}}{\rm{.5}}\;{\rm{nS}} $ ; (c)$ {g_{{\text{syn-e}}}} = {\rm{5}}{\rm{.0}}\;{\rm{nS}} $ ; (d)$ {g_{{\text{syn-e}}}} = {\rm{18}}{\rm{.0}}\;{\rm{nS}} $ .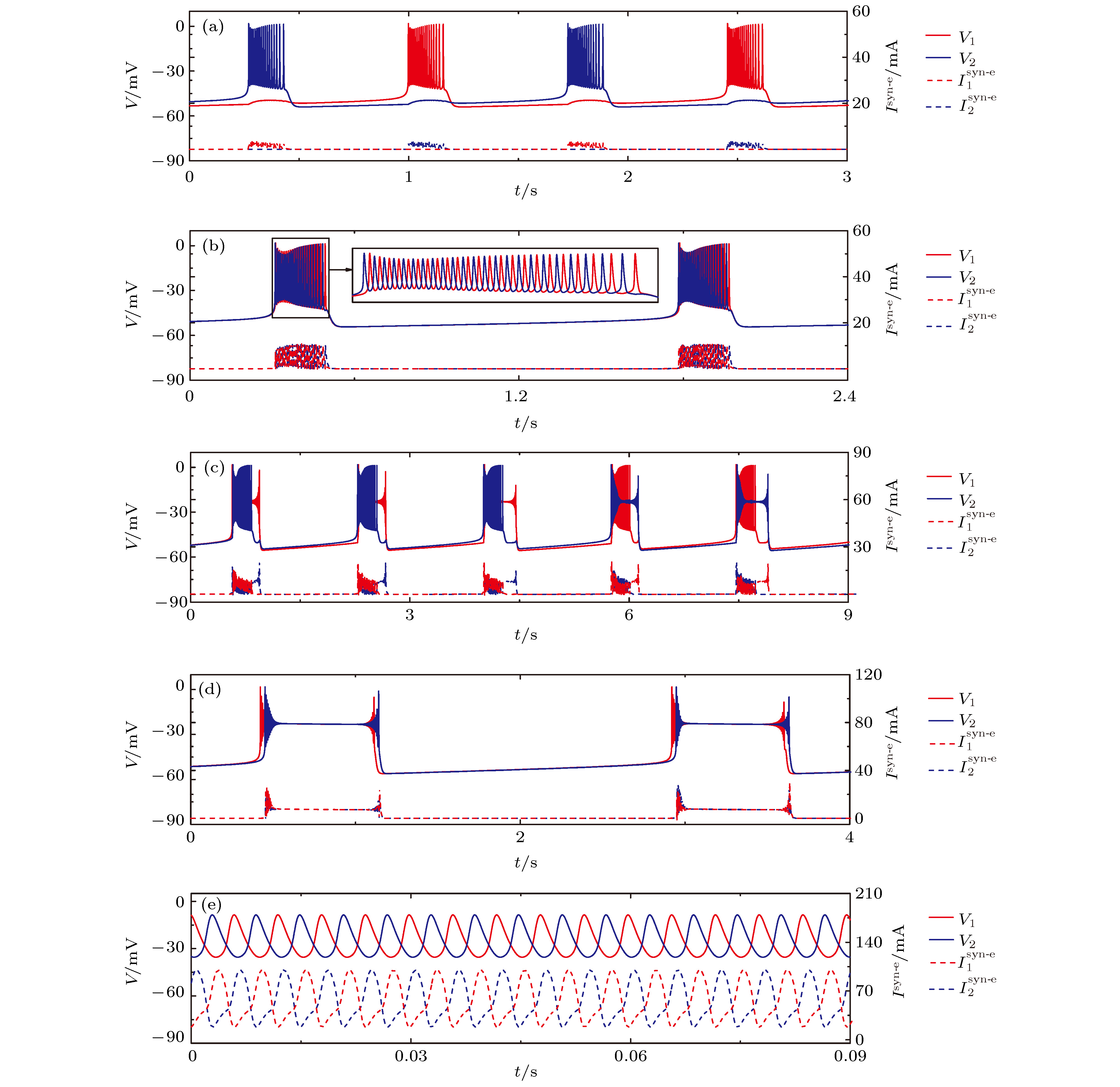
图 7 初值不同时, 不同耦合强度下神经元1(红)和2(蓝)的膜电位V(上)及耦合电流
${I^{{\text{syn-e}}}}$ (下), 插图是局部放大 (a)$g_\text{syn-e}$ = 0.35 nS; (b)$g_\text{syn-e}$ = 1.5 nS (c)$g_\text{syn-e}$ = 2.5 nS; (d)$g_\text{syn-e}$ = 5.0 nS; (e)$g_\text{syn-e}$ = 18.0 nSFig. 7. Membrane potential V (top) and coupling current
${I^{{\text{syn-e}}}}$ (low) of neurons 1 (red) and 2 (blue) with different initial values at different$g_\text{syn-e}$ (Insert figure: the enlargement of bursting): (a)$g_\text{syn-e}$ = 0.35 nS; (b)$g_\text{syn-e}$ = 1.5 nS; (c)$g_\text{syn-e}$ = 2.5 nS; (d)$g_\text{syn-e}$ = 5.0 nS; (e)$g_\text{syn-e}$ = 18.0 nS.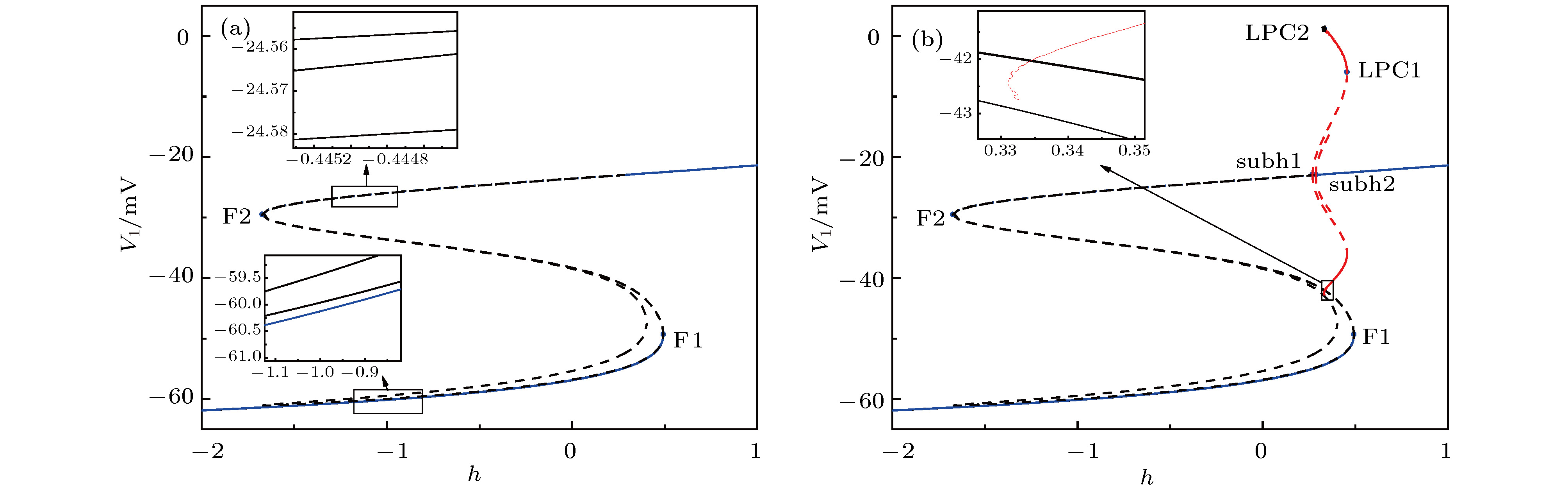
图 8 当
$g_\text{syn-e}$ = 1.5 nS时, 两耦合神经元的快子系统的分岔, 插图是局部放大 (a)平衡点分岔; (b)平衡点分岔和极限环的分岔Fig. 8. Bifurcations of the fast-subsystem of the two coupled neurons with respect to h when
$g_\text{syn-e}$ = 1.5 nS (Insert figure: the enlargement): (a) Equilibrium points; (b) equilibrium points and limit cycle.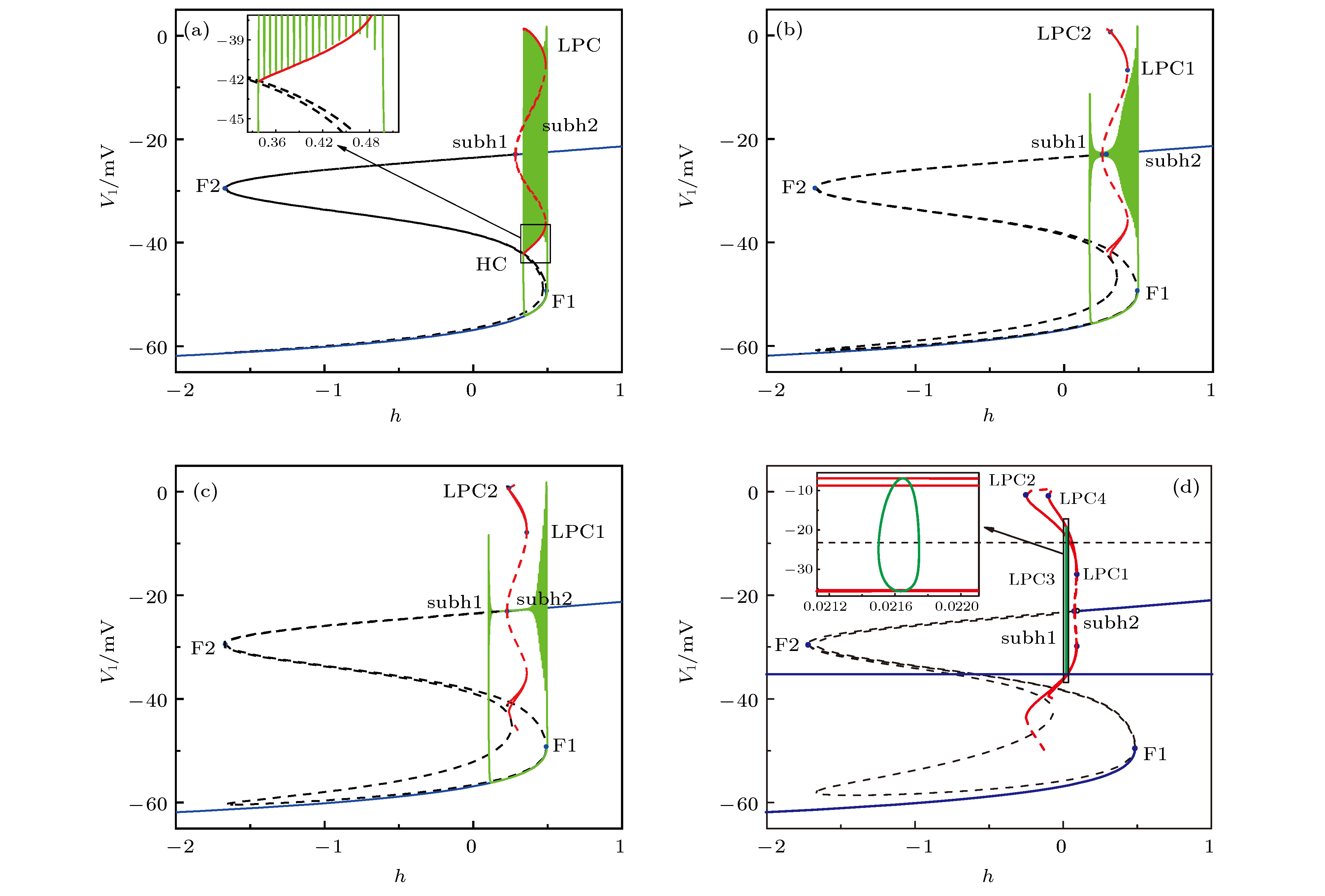
图 9 初值相同时, 神经元1在不同耦合强度下簇放电模式的快慢变量分离, 插图是局部放大 (a)
$g_\text{syn-e}$ = 0.35 nS; (b)$g_\text{syn-e}$ = 2.5 nS; (c)$g_\text{syn-e}$ = 5.0 nS; (d)$g_\text{syn-e}$ = 18.0 nSFig. 9. The fast-slow variable dissection of neuron 1 for different initial values at different
$g_\text{syn-e}$ values (Insert figure: the enlargement): (a)$g_\text{syn-e}$ = 0.35 nS; (b)$g_\text{syn-e}$ = 2.5 nS; (c)$g_\text{syn-e}$ = 5.0 nS; (d)$g_\text{syn-e}$ = 18.0 nS.
图 10 初值不同时, 神经元1在不同耦合强度下簇放电模式的快慢变量分离, 插图是局部放大 (a)
${g_{{\rm{syn - e}}}}$ = 0.35 nS; (b)$g_\text{syn-e}$ = 1.5 nS; (c)和(d)$g_\text{syn-e}$ = 2.5 nS; (e)$g_\text{syn-e}$ = 5.0 nS; (f)$g_\text{syn-e}$ = 18.0 nSFig. 10. The fast-slow variable dissection of neuron 1 for different initial values at different
$g_\text{syn-e}$ values (Insert figure: the enlargement): (a)$g_\text{syn-e}$ = 0.35 nS; (b)$g_\text{syn-e}$ = 1.5 nS; (c) and (d)$g_\text{syn-e}$ = 2.5 nS; (e)$g_\text{syn-e}$ = 5.0 nS; (f)$g_\text{syn-e}$ = 18.0 nS.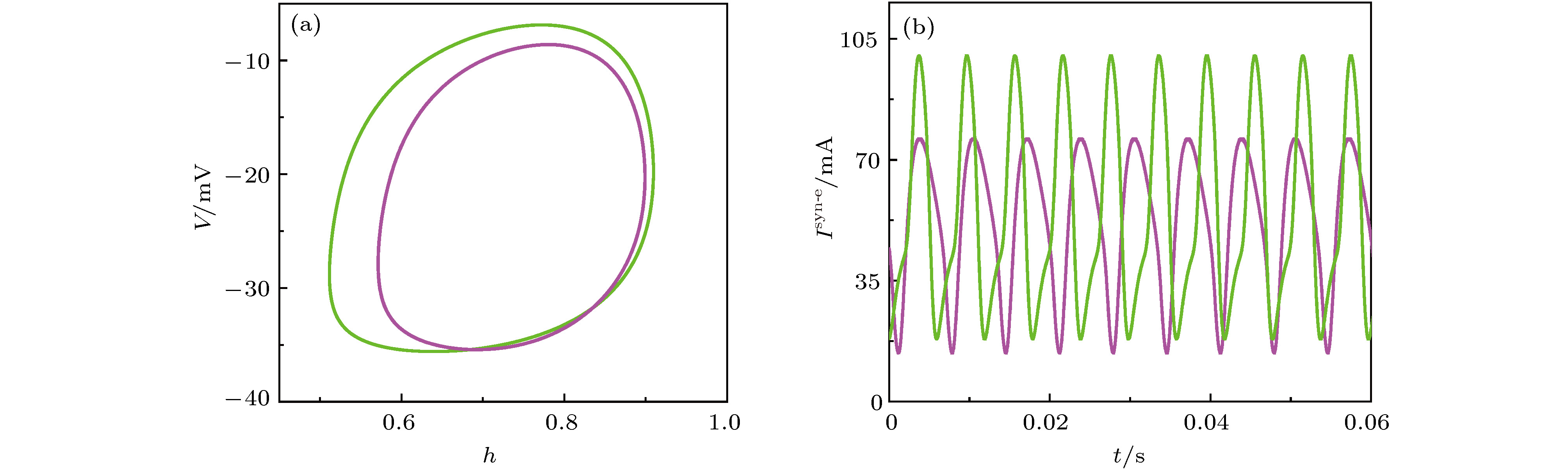
图 11 反相同步(紫色)和同相同步(绿色)周期1峰放电节律 (a) (h, V1)相平面上的相轨迹图; (b)耦合电流随时间t的变化
Fig. 11. The anti-phase (purple) and in-phase (green) period-1 spiking: (a) The V-h trajectory; (b) coupling current.
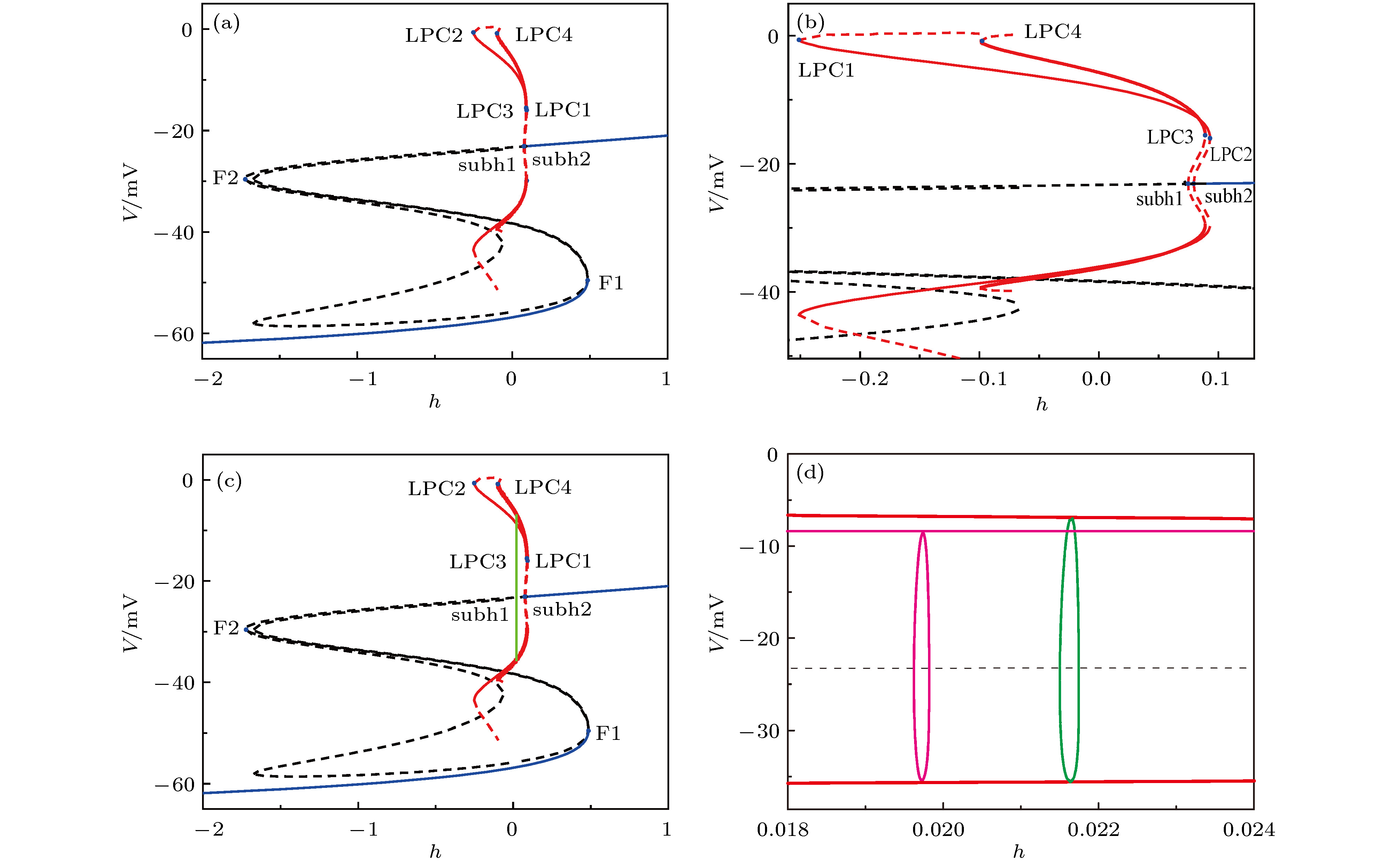
图 12 (a)快子系统的平衡点和极限环的分岔; (b)图(a)中极限环分岔处的放大; (c)反相同步(紫色)和同相同步(绿色)周期1峰放电的快慢变量分离; (d)图(c)中反向同步(紫色)和同向(绿色)同步周期1峰放电的放大
Fig. 12. (a) Bifurcations of equilibrium points and limit cycle of the fast-subsystem; (b) enlargement of (a); (c) fast-slow variable dissection of anti-phase (purple) and in-phase (green) period-1 spiking; (d) enlargement of anti-phase (purple) and in-phase (green) period-1 spiking in Fig. (c).
表 1 理论模型中的参数值
Table 1. Parameter values used in the theoretical model.
参数 参数值 参数 参数值 参数 参数值 参数 参数值 C 21 pF $ {\sigma _{ {\rm{m_p} }} } $ –6 mV $ {g_{ {\rm{Nap} }} } $ 2.8 nS ${E_{{\rm{Na}}}}$ 50 mV $ {\theta _{ {\rm{m_p} }} } $ –40 mV ${\sigma _{\rm{m}}}$ –5 mV ${g_{{\rm{Na}}}}$ 28 nS ${E_{\rm{K}}}$ –85 mV ${\theta _{\rm{m}}}$ –34 mV $\sigma {}_{\rm{h}}$ 6 mV ${g_{\rm{L}}}$ 2.8 nS ${E_{\rm{L}}}$ –65 mV ${\theta _{\rm{h}}}$ –48 mV ${\sigma _{\rm{n}}}$ –4 mV ${g_{ {\text{tonic-e} } } }$ 0.4 nS ${\bar \tau _{\rm{h}}}$ 10000 ms ${\theta _{\rm{n}}}$ –29 mV ${\sigma _{\rm{s}}}$ –5 mV ${\varepsilon _{}}$ 6 ${\bar \tau _{\rm{n}}}$ 5 ms $\theta {}_{\rm{s}}$ –10 mV ${\alpha _{\rm{s}}}$ –5 mV  下载: 导出CSV
下载: 导出CSV
表 2 不同
${g_{\rm{K}}}$ 下快子系统中关键点的慢变量h的值Table 2. The values of slow variable h of the bifurcation or key points at different
${g_{\rm{K}}}$ values.关键点 h的值 F1 F2 subh HC LPC 共存区域 $ {g_{\rm{K} }} = 7.1\;{\rm{nS}} $ 0.4928 –1.6780 0.2128 0.3265 0.4308 [0.3265, 0.4308] $ {g_{\rm{K} }} = 7.8\;{\rm{nS}} $ 0.4928 –1.6680 0.2858 0.3476 0.4973 [0.3476, 0.4928] $ {g_{\rm{K} }} = 10.0 \;{\rm{nS}} $ 0.4928 –1.6390 0.5072 0.3941 0.7025 [0.3941, 0.4928] $ {g_{\rm{K} }} = 25.0 \;{\rm{nS}} $ 0.4928 –1.4800 1.7880 0.4849 1.9240 [0.4849, 0.4928]
下载: 导出CSV
表 3 不同
${g_{{\rm{syn\text-e}}}}$ 下快子系统中关键点的慢变量h的值Table 3. The slow variable h values of the bifurcation or key points at different
${g_{{\rm{syn\text-e}}}}$ values.关键点 h的值 $g_\text{syn-e}$ = 0.35 nS $g_\text{syn-e}$ = 2.5 nS $g_\text{syn-e}$ = 5.0 nS $g_\text{syn-e}$ = 18.0 nS F1 0.4874 0.4918 0.4908 0.4856 F2 –1.6695 –1.6759 –1.6685 –1.7212 subh1 0.2817 0.2565 0.2259 0.0746 subh2 0.2858 0.2852 0.2274 0.0794 LPC1 0.4927 0.4273 0.3598 0.0960 LPC2 \ 0.3103 0.2406 –0.2504 LPC3 \ \ \ 0.0890 LPC4 \ \ \ –0.099 HC 0.3398 \ \ \ 共存区域 [0.3398, 0.4927] [0.3103, 0.4273] [0.2406, 0.3598] [0.0960, 0.250]和[0.0890, 0.099]
下载: 导出CSV
-
[1] Braun H A, Wissing H, Schäfer K, Hirsch M C 1994 Nature 367 270
Google Scholar
[2] Gu H G, Pan B B 2015 Nonlinear Dyn. 81 2107
Google Scholar
[3] 谢勇, 程建慧 2017 物理学报 66 090501
Google Scholar
Xie Y, Cheng J H 2017 Acta Phys. Sin. 66 090501
Google Scholar
[4] Sun X J, Perc M, Kurths J, Lu Q S 2018 Chaos 28 106310
Google Scholar
[5] 徐莹, 王春妮, 靳伍银, 马军 2015 物理学报 64 198701
Xu Y, Wang C N, Jin W Y, Ma J 2015 Acta Phys. Sin. 64 198701
[6] 李国芳, 孙晓娟 2017 物理学报 66 240501
Google Scholar
Li G F, Sun X J 2017 Acta Phys. Sin. 66 240501
Google Scholar
[7] Bianchi A L, Denavit-Saubie M, Champagnat J 1995 Physiol. Rev. 75 1
Google Scholar
[8] Cohen M I 1979 Physiol. Rev. 59 1105
Google Scholar
[9] Funk G D, Smith J C, Feldman J L 1995 J. Neurosci. 15 4046
Google Scholar
[10] Richter D W, Ballanyi K, Schwarzacher S 1992 Curr. Opin. Neurobiol. 2 788
Google Scholar
[11] 严亨秀, 张承武, 郑煜 2004 生理学报 56 665
Google Scholar
Yan H X, Zhang C W, Zheng Y 2004 Acta Physiol. Sin. 56 665
Google Scholar
[12] 宋刚 1999 生理科学进展 3 237
Song G 1999 Prog. Physiol. Sci. 3 237
[13] Smith J C, Ellenberger H H, Ballanyi K, Richter D W, Feldman J L 1991 Science 254 726
Google Scholar
[14] Feldman J L, Negro C A D 2006 Nat. Rev. Neurosci. 7 232
Google Scholar
[15] Smith J C 1997 Neurons, Networks, and Motor Behavior (Cambridge, MA: MIT Press) p97
[16] Johnson S M, Smith J C, Funk G D, Feldman J L 1994 J. Neurophysiol. 72 2598
Google Scholar
[17] Ramirez J M, Richter D W 1996 Curr. Opin. Neurobiol. 6 817
Google Scholar
[18] Rekling J C, Feldman J L 1998 Annu. Rev. Physiol. 60 385
Google Scholar
[19] Koshiya N, Smith J C 1998 28th Annual Meeting of the Society for Neuroscience Los Angeles, California, USA, November 7-12, 1998 p531
[20] Koshiya N, Smith J C 1999 Nature 400 360
Google Scholar
[21] Negro C A D, Morgado V C, Hayes J A, Mackay D D, Pace R W, Crowder E A, Feldman J L 2005 J. Neurosci. 25 446
Google Scholar
[22] Smith J C, Butera R J, Koshiya N, Del Negro C, Wilson C G, Johnson S M 2000 Resp. Physiol. 122 131
Google Scholar
[23] Gray P A, Rekling J C, Bocchiaro C M, Feldman J L 1999 Science 286 1566
Google Scholar
[24] Butera R J, Rinzel J, Smith J C 1999 J. Neurophysiol. 82 382
Google Scholar
[25] Dunmyre J R, Negro C A D, Rubin J E 2011 J. Comput. Neurosci. 31 305
Google Scholar
[26] Negro C A D, Johnson S M, Butera R J, Smith J C 2001 J. Neurophysiol. 86 59
Google Scholar
[27] Butera R J, Rinzel J, Smith J C 1999 J. Neurophysiol. 82 398
Google Scholar
[28] Purvis L K, Smith J C, Koizumi H, Butera R J 2007 J. Neurophysiol. 97 1515
Google Scholar
[29] Best J, Borisyuk A, Rubin J E, Terman D, Wechselberger M 2005 SIAM J. Appl. Dyn. Syst. 4 1107
Google Scholar
[30] Rubin J E 2006 Phys. Rev. E 74 021917
Google Scholar
[31] Dunmyre J R, Rubin J E 2010 SIAM J Appl. Dyn. Syst. 9 154
Google Scholar
[32] Guo D D, Lü Z S 2019 Chin. Phys. B 28 110501
Google Scholar
[33] Rybak I A, Molkov Y I, Jasinski P E, Shevtsova N A, Smith J C 2014 Prog. Brain. Res. 209 1
Google Scholar
[34] 张应腾, 熊冬生, 刘深泉 2015 中国医学物理学杂志 32 115
Google Scholar
Zhang Y T, Xiong D S, Liu S Q 2015 Chin. J. Med. Phys. 32 115
Google Scholar
[35] 刘义, 刘深泉 2011 动力学与控制学报 9 257
Google Scholar
Liu Y, Liu S Q 2011 J. Dynam. Cont. 9 257
Google Scholar
[36] Duan L X, Zhai D H, Tang X H 2012 Int. J. Bifurcation Chaos 22 1250114
Google Scholar
[37] Lü Z S, Chen L N, Duan L X 2019 Appl. Math. Model. 67 234
Google Scholar
[38] Lü Z S, Zhang B Z, Duan L X 2017 Cogn. Neurodynamics 11 443
Google Scholar
[39] Wang Z J, Duan L X, Cao Q Y 2018 Chin. Phys. B 27 070502
Google Scholar
[40] Duan L X, Liu J, Chen X, Xiao P C, Zhao Y 2017 Cogn. Neurodynamics 11 91
Google Scholar
[41] Rubin J E, Shevtsova N A, Ermentrout G B, Smith J C, Rybak I A 2009 J. Neurophysiol. 101 2146
Google Scholar
[42] Rubin J E, Bacak B J, Molkov Y I, Shevtsova N A, Smith J C, Rybak I A 2011 J. Comput. Neurosci. 30 607
Google Scholar
[43] 平小方, 刘深泉, 任会霞 2015 动力学与控制学报 13 215
Google Scholar
Ping X F, Liu S Q, Ren H X 2015 J. Dynam. Cont. 13 215
Google Scholar
[44] Belykh I, Shilnikov A 2008 Phys. Rev. Lett. 101 078102
Google Scholar
[45] Wu F Q, Gu H G, Li Y Y 2019 Commun. Nonlinear Sci. Numer. Simul. 79 104924
Google Scholar
[46] 丁学利, 李玉叶 2016 物理学报 65 210502
Google Scholar
Ding X L, Li Y Y 2016 Acta Phys. Sin. 65 210502
Google Scholar
[47] Jia B, Wu Y C, He D, Guo B H, Xue L 2018 Nonlinear Dyn. 93 1599
Google Scholar
[48] Li Y Y, Gu H G, Ding X L 2019 Nonlinear Dyn. 97 2091
Google Scholar
[49] 曹奔, 关利南, 古华光 2018 物理学报 67 240502
Google Scholar
Cao B, Guan L N, Gu H G 2018 Acta Phys. Sin. 67 240502
Google Scholar
[50] Uzuntarla M, Torres J J, Calim A, Barreto E 2019 Neural Networks 110 131
Google Scholar
[51] 埃门创特 B 著 (孝鹏程, 段丽霞, 苏建忠译) 2002 动力系统仿真, 分析与动画—XPPAUT使用指南 (北京: 科学出版社) 第155−167页
Ermentrout B (translated by Xiao P C, Duan L L, Su J Z) 2002 Simulating, Analyzing, and Animating Dynamical systems: A Guide to XPPAUT for Researchers and Students (Beijing: Science Press) p155−167 (in Chinese)
[52] Izhikevich E M 2000 Int. J. Bifurcation Chaos 10 1171
Google Scholar
[53] Hashemi M, Valizadeh A, Azizi Y 2012 Phys. Rev. E 85 021917
Google Scholar
[54] Jia B, Gu H G 2017 Int. J. Bifurcation Chaos 27 1750113
Google Scholar
[55] Zhao Z G, Gu H G 2017 Sci. Rep. 7 6760
Google Scholar
[56] Li J J, Du M M, Wang R, Lei J Z, Wu Y 2016 Int. J. Bifurcation Chaos 26 1650138
Google Scholar











 下载:
下载:
















































































计量
- 文章访问数: 14165
- PDF下载量: 134
- 被引次数: 0
