










-
钾离子通道在神经细胞动作电位复极过程中起着重要作用. 钾离子通道蛋白种类繁多, 钾离子通道允许钾离子特异性穿过细胞膜, 从而维持神经细胞静息电位. 离子通道蛋白的二级结构决定其功能特性, 皮秒尺度内二级结构的波动会对离子通道蛋白的功能, 即离子通过速率有很大的影响. 本文使用分子动力学模拟方法, 模拟施加不同幅值的53.7 THz的太赫兹波对真实KcsA钾通道蛋白二级结构和钾离子通过速率的影响. 研究发现, 在53.7 THz的太赫兹波的作用下, KcsA钾通道蛋白中α螺旋数量减少, β折叠以及卷曲数量增加. 此外, 53.7 THz的太赫兹波能够加速钾离子通过KcsA钾通道. 本文从蛋白质的二级结构分析太赫兹波对钾离子通道蛋白的影响, 为太赫兹波和生物功能分子相互作用之间提供了新的观察角度.Potassium channels play an important role in repolarizing the nerve cell action potentials. There are many types of potassium channel proteins, and potassium channels allow potassium ions to specifically pass through the cell membrane, thereby maintaining the resting potential of nerve cells. In this paper, molecular dynamics simulation method is used to simulate the effects of 53.7 THz terahertz wave with different amplitudes on the secondary structure of KcsA potassium channel protein and the potassium ions rate. It is found in this study that under the action of the 53.7 THz terahertz wave, the number of alpha helices in KcsA potassium channel protein decreases, and the number of beta sheets and the number of coils increase. In addition, the 53.7 THz terahertz wave can accelerate potassium ions through the KcsA potassium channel. In this article, the effects of terahertz waves on potassium channel proteins are analyzed through the secondary structure of proteins, and a new perspective for the interaction between terahertz waves and biological functional molecules is presented as well.
-
Keywords:
- potassium channel /
- protein secondary structure /
- potassium ion rate /
- resonance absorption
[1] Nelson M T, Quayle J M 1995 Am. J. Physiol. 268 C799
 Google Scholar
Google Scholar
[2] Faraci F M, Sobey C G 1996 Clin. Exp. Pharmacol. Physiol. 23 1091
Google Scholar
[3] Orias M 1998 Medicina 58 429
[4] Johnston J, Forsythe I D, Kopp-Scheinpflug C 2010 J. Physiol. 588 3187
Google Scholar
[5] Yellen G 2002 Nature 419 35
Google Scholar
[6] Guan D, Lee J C F, Higgs M H, Spain W J, Foehring R C 2007 J. Neurophys. 97 1931
Google Scholar
[7] Guan D, Armstrong W E, Foehring R C 2013 J. Physiol. 591 4807
Google Scholar
[8] Zhu Z, Cheng C, Chang C, Ren G, Zhang J, Peng Y, Han J, Zhao H 2019 Analyst 144 2504
Google Scholar
[9] 周俊, 刘盛纲 2014 现代应用物理 5 85
Google Scholar
Zhou J, Liu S G 2014 Modern Applied Physics 5 85
Google Scholar
[10] Michele J C, Piero U 2021 Chem. Phys. 155 075102
Google Scholar
[11] Sizov F 2017 SPQEO 20 273
Google Scholar
[12] Li N, Peng D L, Zhang X J, Shu Y S, Zhang F, Jiang L, Song B 2021 Nano Res. 14 40
Google Scholar
[13] Wilmink G J, Grundt J E 2011 J. Infrared Millimeter Terahertz Waves 32 1074
Google Scholar
[14] Bo W F, Guo L H, Yang Y, Ma J L, Wang K C, Tang J C, Wu Z, Zeng B Q, Gong Y B 2020 IEEE Access 8 10305
Google Scholar
[15] Li Y M, Chang C, Zhu Z, Sun L, Fan C H 2021 JACS 143 4311
Google Scholar
[16] Liu X, Qiao Z, Chai Y M, Zhu Z, Wu K J, Ji W L, Li D G, Xiao Y J, Mao L Q, Chang C, Wen Q, Song B, Shu Y S 2021 PNAS 118 e2015685118
Google Scholar
[17] Zhang J X, He Y, Liang S S, Liao X, Li T, Qiao Z, Chang C, Jia H B, Chen X W 2021 Nat. Commun. 12 2730
Google Scholar
[18] Zhu Z, Chen C, Chang C, Song B 2021 ACS Photonics 8 781
Google Scholar
[19] Zhang X X, He M X, Chen Y, Li C, Zhao J W, Wang P F, Peng X 2019 Chin. Phys. B 28 128702
Google Scholar
[20] Alexandrov B S, Rasmussen K Ø, Bishop A R, Usheva A, Rodriguez G 2011 Biomed. Opt. Express 2 2679
Google Scholar
[21] Yamazaki S, Harata M, Ueno Y, Tsubouchi M, Konagaya K, Ogawa Y, Isoyama G, Otani C, Hoshina H 2020 Sci. Rep. 10 9008
Google Scholar
[22] Yamazaki S, Harata M, Idehara T, Konagaya K, Yokoyama G, Hoshina H, Ogawa Y 2018 Sci. Rep. 8 9990
Google Scholar
[23] Wu K J, Qi C H, Zhu Z, Wang C L, Song B, Chang C 2020 J. Phys. Chem. Lett. 11 7002
Google Scholar
[24] Takehiro T, Reiko S, Shiho T, Ken-Ichiro K, Hideki H 2020 Opt. Lett. 45 6078
Google Scholar
[25] Jorgensen W L, Chandrasekhar J, Madura J D, Impey R W, Klein M L 1983 J. Chem. Phys. 79 926
Google Scholar
[26] Biggin P C, Smith G R, Shrivastava I, Choe S, Sansom M S P 2001 BBA-Biomemberanes 1510 1
Google Scholar
[27] Berendsen H J C, Spoel D V D, Drunen R V 1995 Comput. Phys. Commun. 91 43
Google Scholar
[28] Parker M J, Sessions R B, Badcoe I G, Clarke A R 1996 Fold Des. 1 145
Google Scholar
[29] Zhou H X, Wlodek S T, McCammon J A 1998 PNAS 95 9280
Google Scholar
[30] Barron L D, Hecht L, Wilson G 1997 Biochemistry 36 13143
Google Scholar
[31] Fischer S, Smith J C, Verma C S 2001 J. Phys. Chem. B 105 8050
Google Scholar
[32] Leach A R 2001 Molecular Modelling: Principles and Applications (2nd Ed.) (Harlow: Pearson Education Ltd.) pp20−30
[33] Rath A, Johnson R M, Deber C M 2007 Pept. Sci. 88 217
Google Scholar
[34] Moore D T, Berger B W, DeGrado W F 2008 Structure 16 991
Google Scholar
[35] Matthews E E, Zoonens M, Engelman D M 2006 Cell 127 447
Google Scholar
-
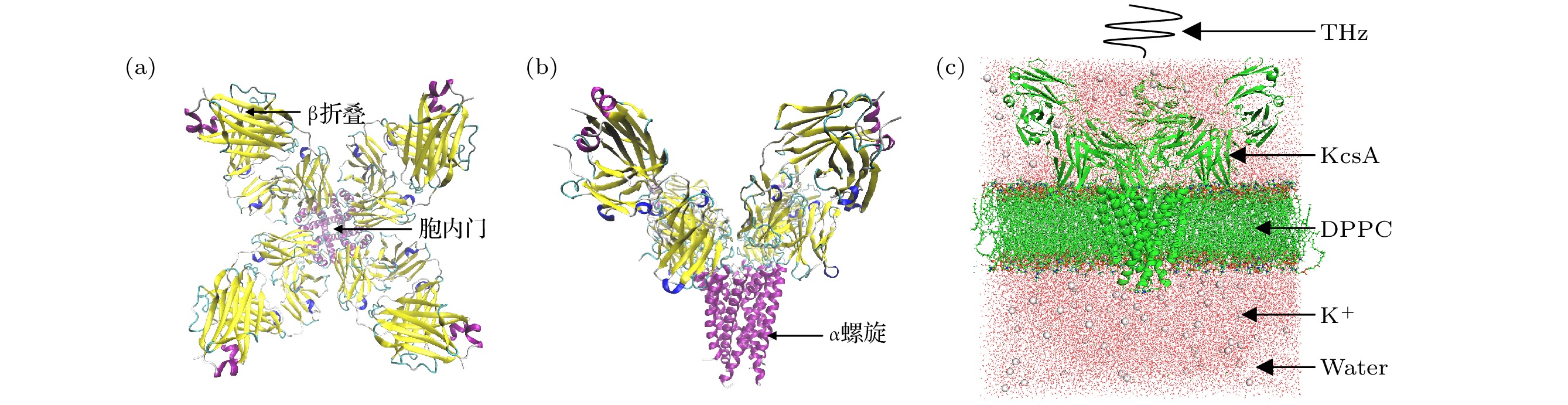
图 1 (a) KcsA四聚体结构(从上往下); (b) KcsA四聚体结构(从前往后); (c)钾离子通道模型
Fig. 1. (a) KcsA tetramer structure (from top to bottom); (b) KcsA tetramer structure (from front to back); (c) potassium channel model.
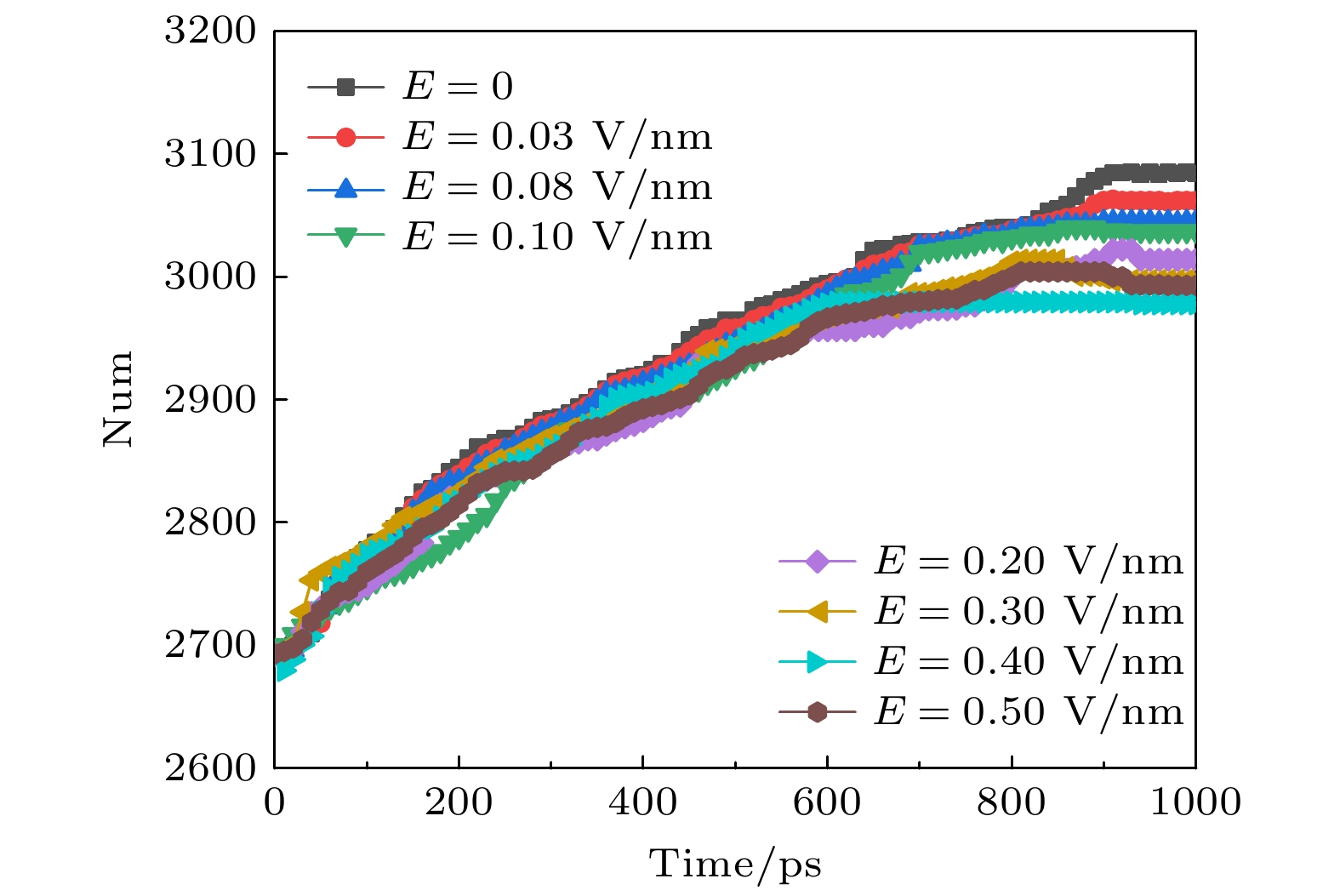
图 2 太赫兹波强度不同时钾离子通道蛋白中氢键的变化
Fig. 2. Changes of hydrogen bonds in potassium channel proteins at different terahertz intensities.
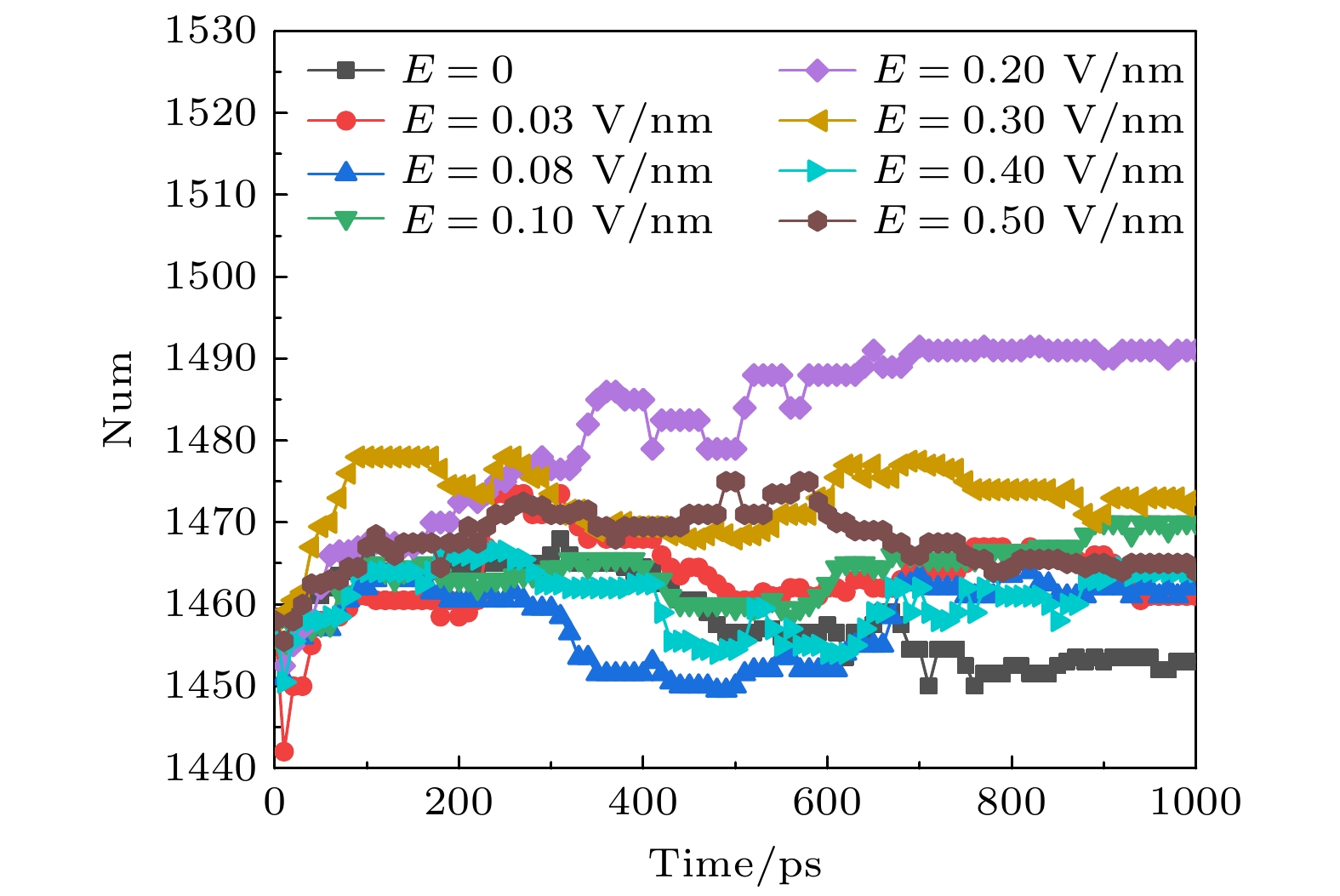
图 3 太赫兹波强度不同时钾离子通道蛋白中二级结构数目的变化
Fig. 3. Changes in the number of secondary structures in potassium channel proteins under different terahertz intensities.
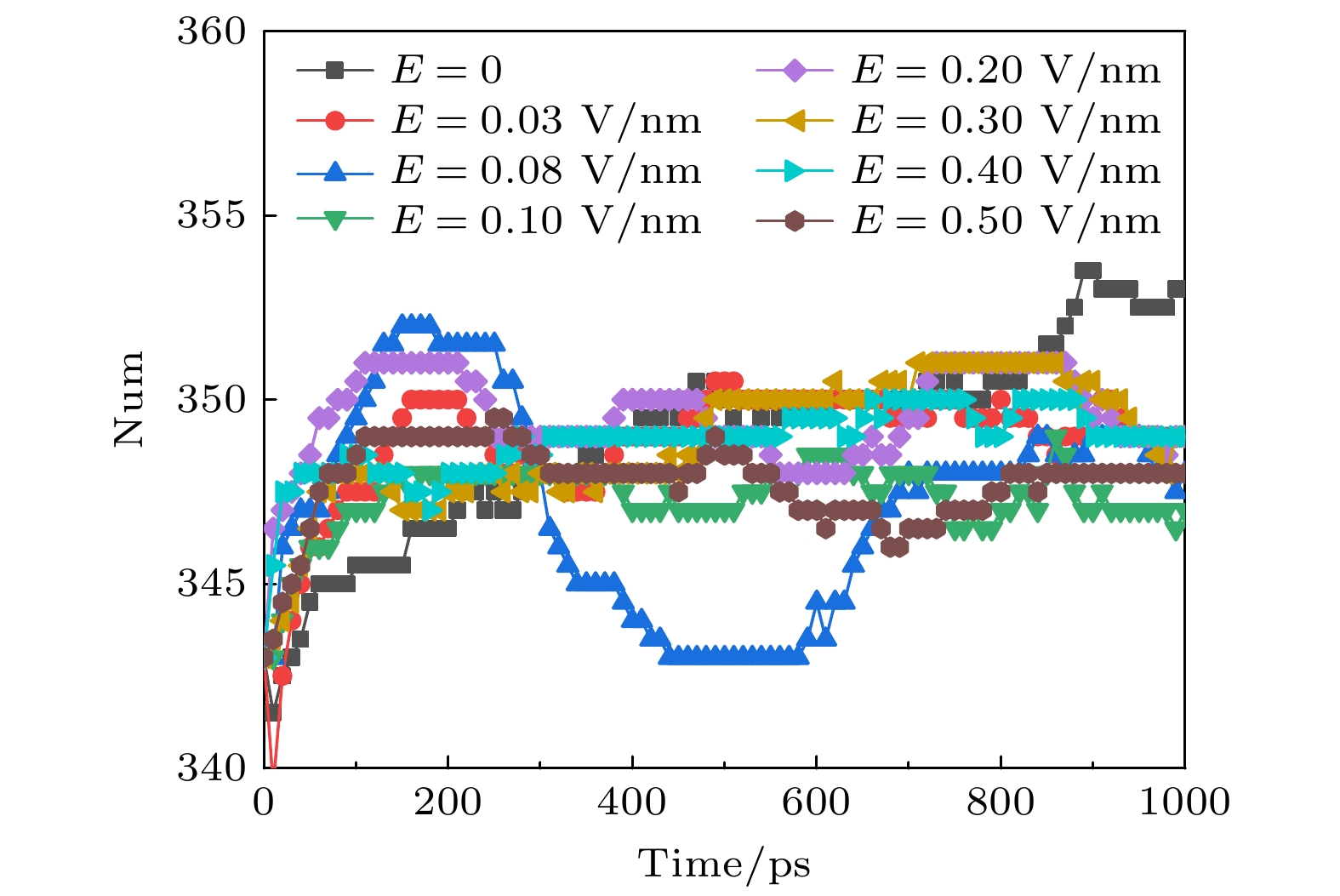
图 4 太赫兹波强度不同时钾离子通道蛋白中α螺旋数目的变化
Fig. 4. Changes in the number of α-helices in potassium channel protein under different terahertz intensities.
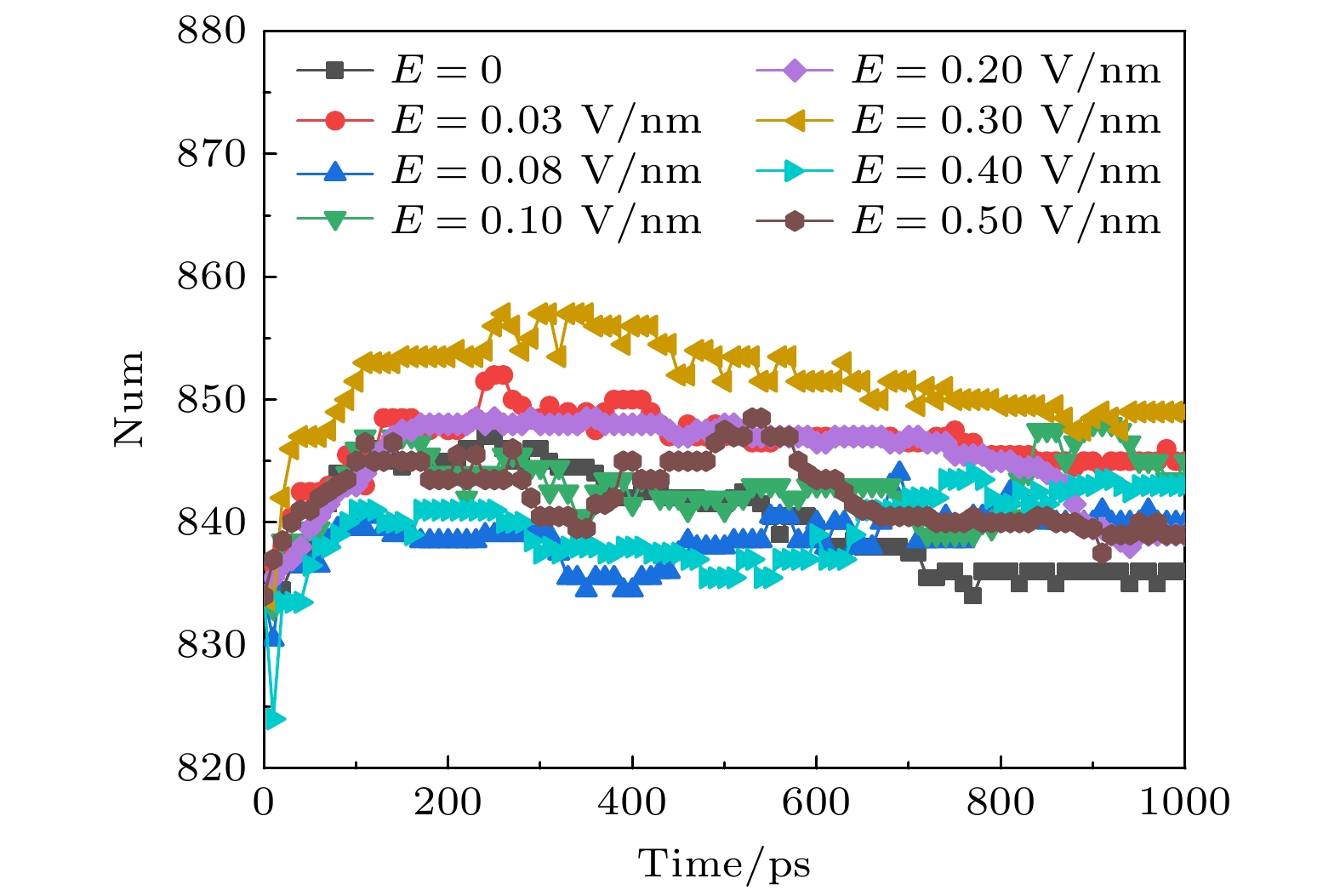
图 5 太赫兹波强度不同时钾离子通道蛋白中β折叠数目的变化
Fig. 5. Changes in the number of β-sheet in potassium channel protein under different terahertz intensities.

图 6 太赫兹波强度不同时钾离子通道蛋白中卷曲数目的变化
Fig. 6. Changes in the number of coil in potassium channel protein under different terahertz intensities.
-
[1] Nelson M T, Quayle J M 1995 Am. J. Physiol. 268 C799
Google Scholar
[2] Faraci F M, Sobey C G 1996 Clin. Exp. Pharmacol. Physiol. 23 1091
Google Scholar
[3] Orias M 1998 Medicina 58 429
[4] Johnston J, Forsythe I D, Kopp-Scheinpflug C 2010 J. Physiol. 588 3187
Google Scholar
[5] Yellen G 2002 Nature 419 35
Google Scholar
[6] Guan D, Lee J C F, Higgs M H, Spain W J, Foehring R C 2007 J. Neurophys. 97 1931
Google Scholar
[7] Guan D, Armstrong W E, Foehring R C 2013 J. Physiol. 591 4807
Google Scholar
[8] Zhu Z, Cheng C, Chang C, Ren G, Zhang J, Peng Y, Han J, Zhao H 2019 Analyst 144 2504
Google Scholar
[9] 周俊, 刘盛纲 2014 现代应用物理 5 85
Google Scholar
Zhou J, Liu S G 2014 Modern Applied Physics 5 85
Google Scholar
[10] Michele J C, Piero U 2021 Chem. Phys. 155 075102
Google Scholar
[11] Sizov F 2017 SPQEO 20 273
Google Scholar
[12] Li N, Peng D L, Zhang X J, Shu Y S, Zhang F, Jiang L, Song B 2021 Nano Res. 14 40
Google Scholar
[13] Wilmink G J, Grundt J E 2011 J. Infrared Millimeter Terahertz Waves 32 1074
Google Scholar
[14] Bo W F, Guo L H, Yang Y, Ma J L, Wang K C, Tang J C, Wu Z, Zeng B Q, Gong Y B 2020 IEEE Access 8 10305
Google Scholar
[15] Li Y M, Chang C, Zhu Z, Sun L, Fan C H 2021 JACS 143 4311
Google Scholar
[16] Liu X, Qiao Z, Chai Y M, Zhu Z, Wu K J, Ji W L, Li D G, Xiao Y J, Mao L Q, Chang C, Wen Q, Song B, Shu Y S 2021 PNAS 118 e2015685118
Google Scholar
[17] Zhang J X, He Y, Liang S S, Liao X, Li T, Qiao Z, Chang C, Jia H B, Chen X W 2021 Nat. Commun. 12 2730
Google Scholar
[18] Zhu Z, Chen C, Chang C, Song B 2021 ACS Photonics 8 781
Google Scholar
[19] Zhang X X, He M X, Chen Y, Li C, Zhao J W, Wang P F, Peng X 2019 Chin. Phys. B 28 128702
Google Scholar
[20] Alexandrov B S, Rasmussen K Ø, Bishop A R, Usheva A, Rodriguez G 2011 Biomed. Opt. Express 2 2679
Google Scholar
[21] Yamazaki S, Harata M, Ueno Y, Tsubouchi M, Konagaya K, Ogawa Y, Isoyama G, Otani C, Hoshina H 2020 Sci. Rep. 10 9008
Google Scholar
[22] Yamazaki S, Harata M, Idehara T, Konagaya K, Yokoyama G, Hoshina H, Ogawa Y 2018 Sci. Rep. 8 9990
Google Scholar
[23] Wu K J, Qi C H, Zhu Z, Wang C L, Song B, Chang C 2020 J. Phys. Chem. Lett. 11 7002
Google Scholar
[24] Takehiro T, Reiko S, Shiho T, Ken-Ichiro K, Hideki H 2020 Opt. Lett. 45 6078
Google Scholar
[25] Jorgensen W L, Chandrasekhar J, Madura J D, Impey R W, Klein M L 1983 J. Chem. Phys. 79 926
Google Scholar
[26] Biggin P C, Smith G R, Shrivastava I, Choe S, Sansom M S P 2001 BBA-Biomemberanes 1510 1
Google Scholar
[27] Berendsen H J C, Spoel D V D, Drunen R V 1995 Comput. Phys. Commun. 91 43
Google Scholar
[28] Parker M J, Sessions R B, Badcoe I G, Clarke A R 1996 Fold Des. 1 145
Google Scholar
[29] Zhou H X, Wlodek S T, McCammon J A 1998 PNAS 95 9280
Google Scholar
[30] Barron L D, Hecht L, Wilson G 1997 Biochemistry 36 13143
Google Scholar
[31] Fischer S, Smith J C, Verma C S 2001 J. Phys. Chem. B 105 8050
Google Scholar
[32] Leach A R 2001 Molecular Modelling: Principles and Applications (2nd Ed.) (Harlow: Pearson Education Ltd.) pp20−30
[33] Rath A, Johnson R M, Deber C M 2007 Pept. Sci. 88 217
Google Scholar
[34] Moore D T, Berger B W, DeGrado W F 2008 Structure 16 991
Google Scholar
[35] Matthews E E, Zoonens M, Engelman D M 2006 Cell 127 447
Google Scholar

 下载:
下载:

计量
- 文章访问数: 12539
- PDF下载量: 215
- 被引次数: 0
