










-
针对大口径干涉型红外光谱辐射计, 分析了不平行于主光轴的入射光, 对理想仪器线型函数的影响. 本文系统介绍了对仪器线型函数产生影响的截断效应、有限视场效应、离轴效应和离焦效应, 并通过HITRAN数据库对理想的水汽吸收光谱进行仿真, 建立了仪器线型函数误差与光谱畸变的定量关系. 根据仿真结果, 提出了因子权重校正算法. 利用水汽吸收的仿真数据对因子权重校正算法进行验证, 光谱漂移从0.51 cm–1降低到0.01 cm–1以下. 通过自研干涉型红外光谱辐射计对标准黑体的观测实验, 验证因子权重校正算法的准确性, 实测数据的光谱漂移从0.226 cm–1降低到0.012 cm–1, 校正后的光谱数据更为准确.
-
关键词:
- 遥感 /
- 干涉型红外光谱辐射计 /
- 仪器线型函数 /
- 因子权重法
Interferometric infrared spectral radiometer has high luminous flux and large passing aperture, so the spectral data collected by the instrument are the convolution of the target spectral data and the instrument line shape (ILS). The main factors affecting the ILS include truncation effect, finite field of view effect, off-axis effect, defocus effect and relative position of detector and so on. In this paper the truncation effect, finite field of view effect, off-axis effect and defocus effect on ILS are expounded. These ILS errors all cause the ideal spectrum to drift towards the low wave number and widen. In this paper, the ideal absorption spectrum of water vapor is simulated by the line-by-line integration, through using MATLAB software and combining with the data of water vapor spectrum in HITRAN database, and the quantitative relationship between the error of instrument linear function and spectral distortion is established. According to the simulation results, a factor weight correction algorithm is proposed. It is believed that with this method, the error spectrum caused by ILS has the same optical properties as the ideal spectrum, but the optical path difference is different. Therefore, the coefficient matrix H can be constructed to establish the quantitative relationship between the error spectrum and the ideal spectrum, and the error spectrum can be corrected by using Landweber iterative algorithm. In this paper, the simulation data of water vapor absorption are used to verify the factor weight correction algorithm, and the spectral drift decreases from 0.51 cm–1 to less than 0.01 cm–1. The accuracy of the factor weight correction algorithm is verified by the experimental observation of the standard black body with self-developed interferometric infrared spectral radiometer. The spectral drift of the measured data decreases from 0.226 cm–1 to 0.012 cm–1, and the corrected spectral data are more accurate.-
Keywords:
- remote sensing /
- infrared spectrometer /
- instrument line shape /
- weight factor method
[1] 刘文清, 陈臻懿, 刘建国, 谢品华, 张天舒, 阚瑞峰, 徐亮 2018 中国环境监测 34 1
 Google Scholar
Google Scholar
Liu W Q, Chen Z R, Liu J G, Xie P H, Zhang T S, Kan R F, Xu L 2018 Envir. Monitor. China 34 1
Google Scholar
[2] Xiao H K, Levine S P, Nowak J, Puskar M, Spear R C 1993 Am. Ind. Hyg. Assoc. J. 54 545
Google Scholar
[3] Rehault J, Borrego-Varillas R, Oriana A, Manzoni C, Hauri C P, Helbing J, Cerullo G 2017 Opt. Express 25 4403
Google Scholar
[4] 徐亮, 刘建国, 高闽光, 陆亦怀, 刘文清, 张天舒, 魏秀丽, 赵雪松, 朱军, 陈华 2007 大气与环境光学学报 2 60
Google Scholar
Xu L, Liu J G, Gao M G, Lu Y H, Liu W Q, Zhang T S, Wei X L, Zhao X S, Zhu J, Chen H 2007 J. Atmosph. Environ. Opt. 2 60
Google Scholar
[5] 金岭, 徐亮, 高闽光, 童晶晶, 程巳阳, 李相贤 2013 大气与环境光学学报 8 416
Google Scholar
Jin L, Xu L, Gao M G, Dong J J, Cheng S Y, Li X X 2013 J. Atmosph. Environ. Opt. 8 416
Google Scholar
[6] Shen X C, Ye S B, Xu L, Hu R, Jin L, Xu H Y, Liu J G, Liu W Q 2018 Appl. Opt. 57 5794
Google Scholar
[7] Hase F, Blumenstock T, Paton-Walsh C 1999 Appl. Opt. 38 3417
Google Scholar
[8] Wiacek A, Taylor J R, Strong K, Saari R, Kerzenmacher T E, Jones N B, Griffith D W T 2007 J. Atmos. Ocean. Technol. 24 432
Google Scholar
[9] Han Y, Revercomb H, Cromp M, et al. 2013 J Geophys. Res. Atmos. 118 12734
Google Scholar
[10] 张磊, 杨敏珠, 邹曜璞, 韩昌佩 2015 光学精密工程 23 3322
Google Scholar
Zhang L, Yang M Z, Zou Y P, Han C P 2015 Opt. Prec. Eng. 23 3322
Google Scholar
[11] Xu L, Wang J, Liu J G, Gao M G, Lu Y H, Liu W Q, Wei X L, Zhang T S, Chen 徐H, Liu Z M 2007 J. Atmosph. Environ. Opt. 2 218
Google Scholar
徐亮, 王君, 刘建国, 高闽光, 陆亦怀, 刘文清, 魏秀丽, 张天舒, 陈华, 刘志明 2007 大气与环境光学学报 2 218
Google Scholar
[12] 熊伟, 施海亮, 俞能海 2015 光谱学与光谱分析 35 267
Google Scholar
Xiong W, Shi H L, Yu N H 2015 Spectrosc. Spect. Anal. 35 267
Google Scholar
[13] Saarinen P, Kauppinen J 1992 Appl. Opt. 31 2353
Google Scholar
[14] Salonen K I, Salomaa I K, Kauppinen J K 1995 Appl. Opt. 34 1190
Google Scholar
[15] 冯明春, 徐亮, 刘文清, 刘建国, 高闽光, 魏秀丽 2016 物理学报 65 168
Google Scholar
Feng M C, Xu L, Liu W Q, Liu J G, Gao M G, Wei X L 2016 Acta Phys. Sin. 65 168
Google Scholar
[16] 冯明春, 刘文清, 徐亮, 高闽光, 魏秀丽, 童晶晶, 李相贤, 陈军 2015 光学学报 35 0423002
Google Scholar
Feng M C, Liu W Q, Xu L, Gao M G, Wei X L, Tong J J, Li X X, Chen J 2015 Act. Opt. Sini. 35 0423002
Google Scholar
[17] Grohs P, Kereta Z, Wiesmann U 2016 Int. J. Wavelets Multiresolut. Inf. Process. 14 1650032
Google Scholar
[18] Dicke R H 1953 Phys. Rev. 89 472
Google Scholar
[19] Penner S S, Kavanagh R W 1953 J. Opt. Soc. Am. 43 385
Google Scholar
[20] He J, Zhang Q G 2013 Optik 124 5245
Google Scholar
[21] 徐亮, 刘建国, 高闽光, 陆亦怀, 刘文清, 魏秀丽, 张天舒, 陈华, 刘志明 2008 光谱学与光谱分析 28 1052
Google Scholar
Xu L, Liu J G, Gao M G, Lu Y H, Liu W Q, Wei X L, Zhang T S, Chen H, Liu Z M 2008 Spectrosc. Spect. Anal. 28 1052
Google Scholar
-

图 1 波数1555 cm–1和1580 cm–1入射光不同动镜运动距离干涉数据和复原光谱
Fig. 1. Wavenumber 1555 cm–1 and 1580 cm–1 interferometric data and reconstructed spectra of different moving distance of incident light.
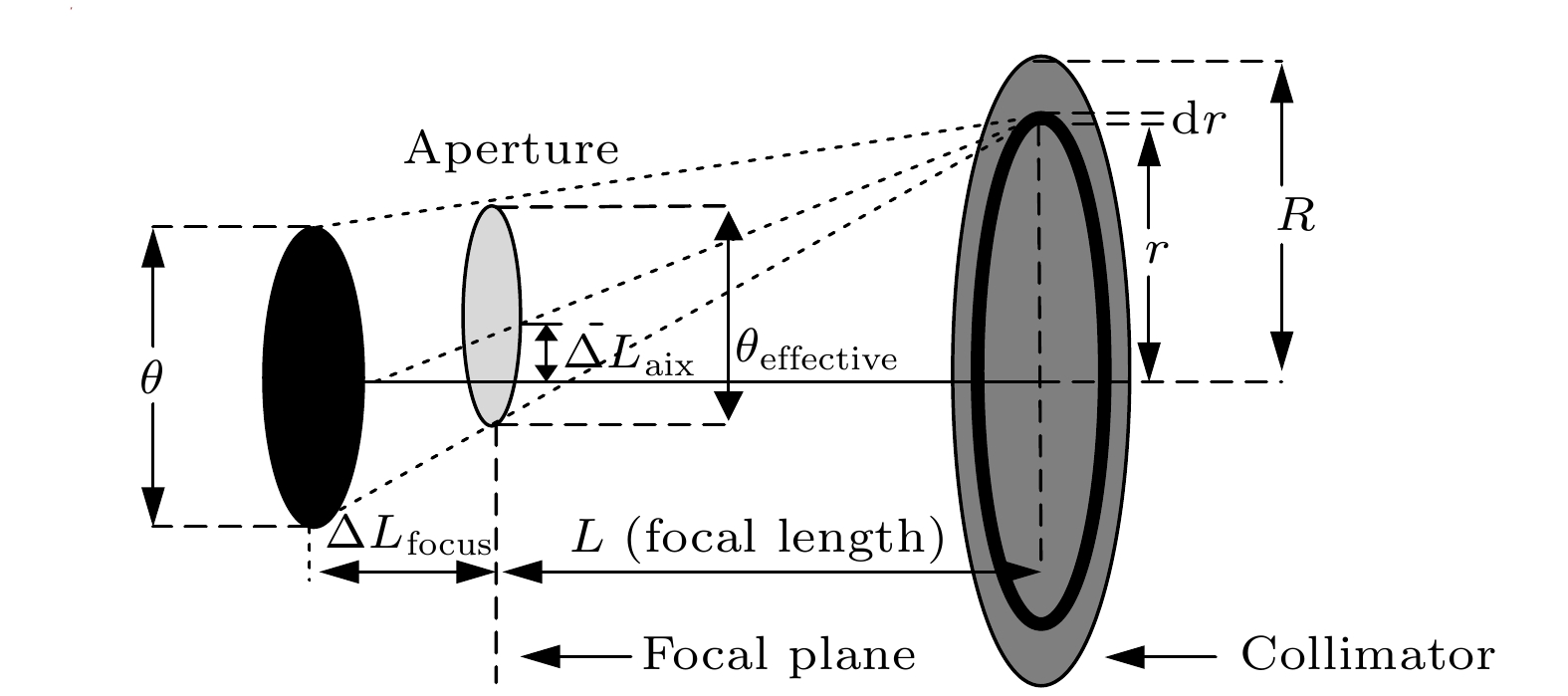
图 3 离焦效应与离轴效应光路示意图
Fig. 3. Diagram of light path of defocus effect and off-axis effect

图 4 理想光谱与误差光谱权重关系示意图
Fig. 4. Diagram of weight relation between ideal spectrum and error spectrum.
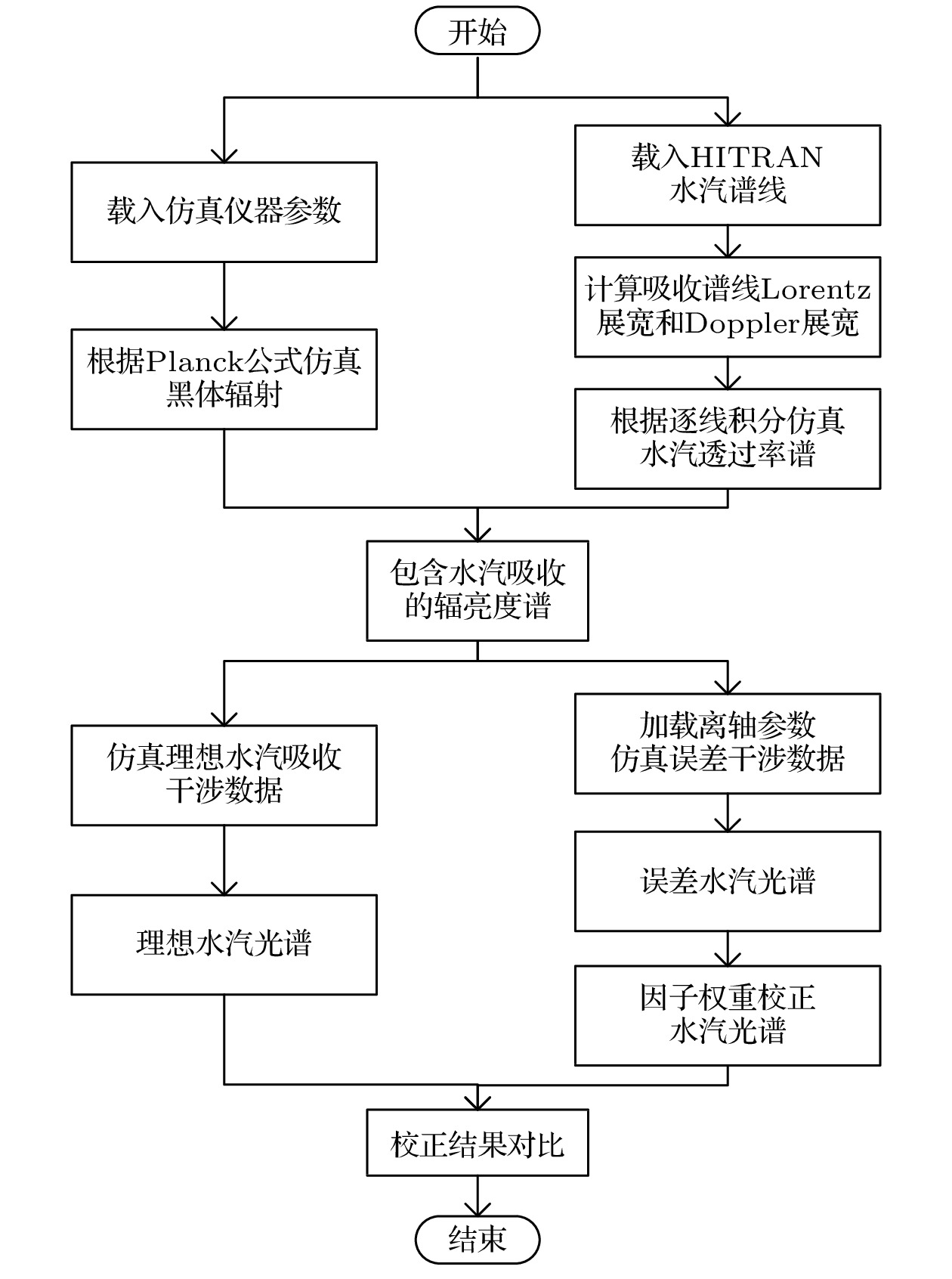
图 5 水汽吸收光谱仿真及校正流程图
Fig. 5. Flow chart of water vapor absorption spectrum simulation and correction.

图 6 (a)水汽透过率谱; (b)包含水汽吸收峰的辐亮度谱
Fig. 6. (a) Water vapor transmittance spectrum; (b) radiance spectra containing water vapor absorption peaks.

图 7 100 ℃时水汽的理想吸收光谱、仪器线型函数误差光谱与校正光谱
Fig. 7. Ideal absorption spectra of water vapor at 100 ℃, error spectra of instrument linear function and correction spectra.
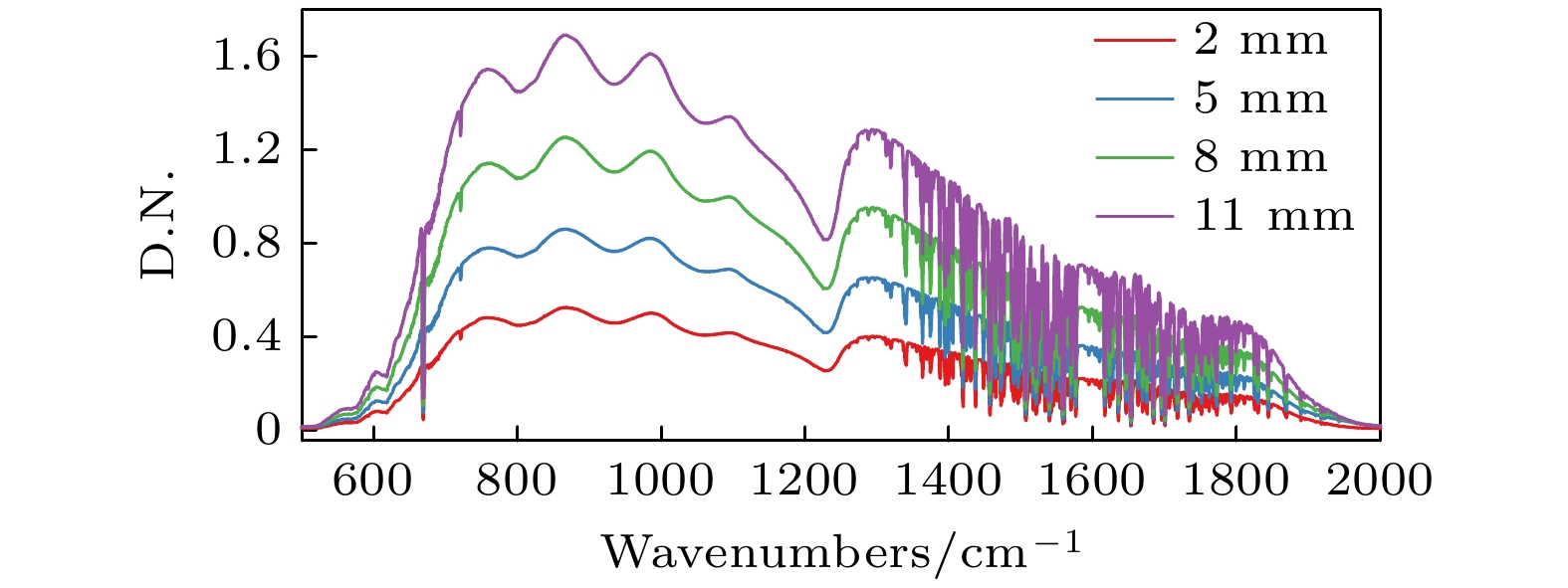
图 8 100 ℃黑体光源不同视场光阑复原光谱图
Fig. 8. Resurrected spectrum of 100 ℃ blackbody source with different field of view by diaphragm.

图 9 100 ℃黑体光源不同视场光阑复原光谱归一化光谱图
Fig. 9. Normalized spectral images of 100 ℃ black-body light source with different field of view apertures.
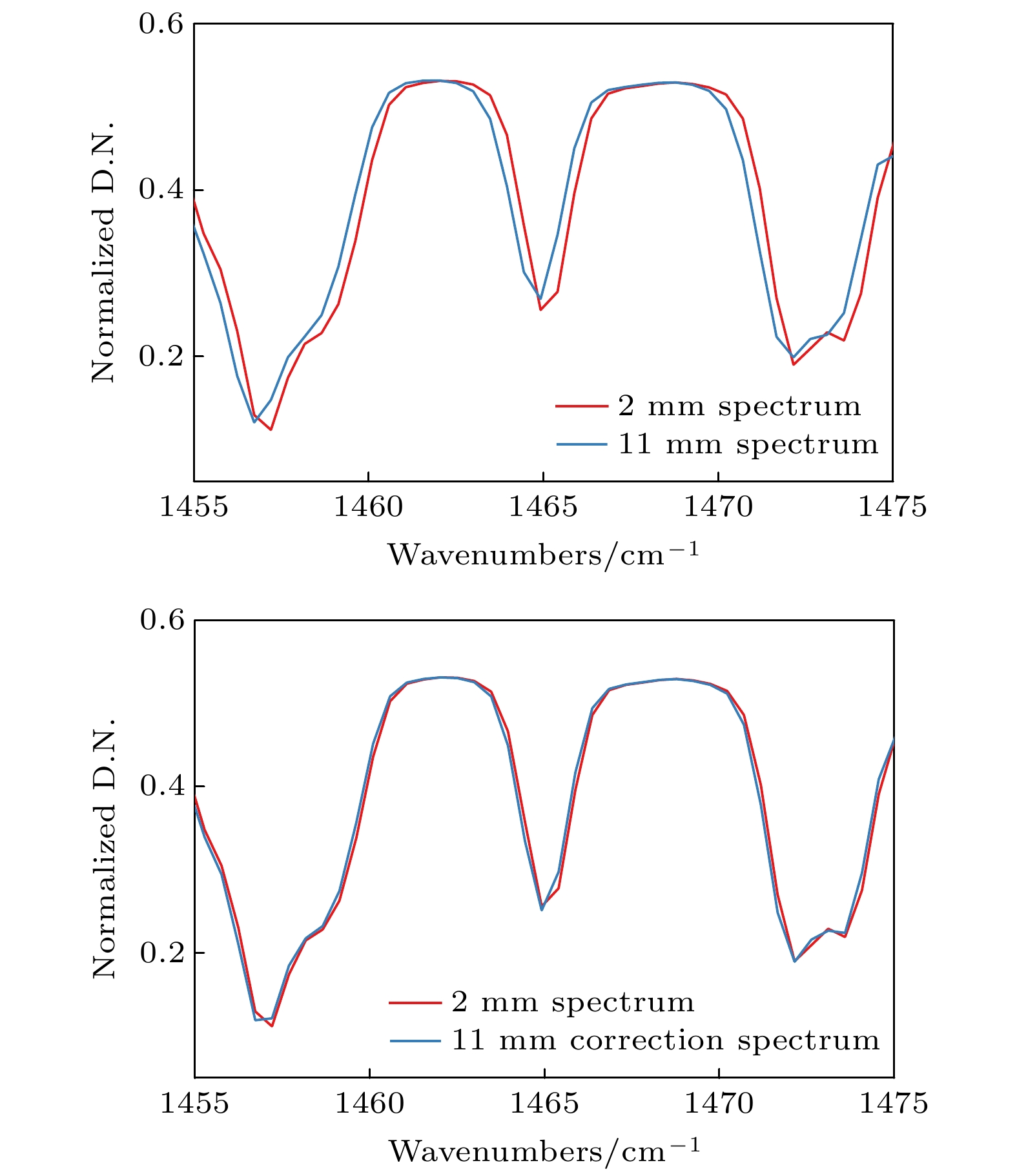
图 10 波数为1465. 2
$ {\mathrm{c}\mathrm{m}}^{-1} $ 的水汽吸收峰校正前后对比Fig. 10. Comparison of water vapor absorption peak with wave number of 1465. 2
$ {\mathrm{c}\mathrm{m}}^{-1} $ before and after correction.表 1 100 ℃黑体光源孔径光阑为11 mm光谱校正前后波数误差对比
Table 1. Comparison of wave number errors before and after spectral correction of 11 mm aperture diaphragm of 100 ℃ blackbody light source.
Wavenumbers /$ {\mathrm{c}\mathrm{m}}^{-1} $ Wavenumber
shift /$ {\mathrm{c}\mathrm{m}}^{-1} $Error-reduction value/$ {\mathrm{c}\mathrm{m}}^{-1} $ Measured spectrum Correction spectrum 1430—1440 0.242 0.018 0.224 1460—1470 0.215 0.008 0.207 1572—1582 0.217 0.010 0.207 1650—1660 0.231 0.012 0.219 Average 0.226 0.012 0.214  下载: 导出CSV
下载: 导出CSV
-
[1] 刘文清, 陈臻懿, 刘建国, 谢品华, 张天舒, 阚瑞峰, 徐亮 2018 中国环境监测 34 1
Google Scholar
Liu W Q, Chen Z R, Liu J G, Xie P H, Zhang T S, Kan R F, Xu L 2018 Envir. Monitor. China 34 1
Google Scholar
[2] Xiao H K, Levine S P, Nowak J, Puskar M, Spear R C 1993 Am. Ind. Hyg. Assoc. J. 54 545
Google Scholar
[3] Rehault J, Borrego-Varillas R, Oriana A, Manzoni C, Hauri C P, Helbing J, Cerullo G 2017 Opt. Express 25 4403
Google Scholar
[4] 徐亮, 刘建国, 高闽光, 陆亦怀, 刘文清, 张天舒, 魏秀丽, 赵雪松, 朱军, 陈华 2007 大气与环境光学学报 2 60
Google Scholar
Xu L, Liu J G, Gao M G, Lu Y H, Liu W Q, Zhang T S, Wei X L, Zhao X S, Zhu J, Chen H 2007 J. Atmosph. Environ. Opt. 2 60
Google Scholar
[5] 金岭, 徐亮, 高闽光, 童晶晶, 程巳阳, 李相贤 2013 大气与环境光学学报 8 416
Google Scholar
Jin L, Xu L, Gao M G, Dong J J, Cheng S Y, Li X X 2013 J. Atmosph. Environ. Opt. 8 416
Google Scholar
[6] Shen X C, Ye S B, Xu L, Hu R, Jin L, Xu H Y, Liu J G, Liu W Q 2018 Appl. Opt. 57 5794
Google Scholar
[7] Hase F, Blumenstock T, Paton-Walsh C 1999 Appl. Opt. 38 3417
Google Scholar
[8] Wiacek A, Taylor J R, Strong K, Saari R, Kerzenmacher T E, Jones N B, Griffith D W T 2007 J. Atmos. Ocean. Technol. 24 432
Google Scholar
[9] Han Y, Revercomb H, Cromp M, et al. 2013 J Geophys. Res. Atmos. 118 12734
Google Scholar
[10] 张磊, 杨敏珠, 邹曜璞, 韩昌佩 2015 光学精密工程 23 3322
Google Scholar
Zhang L, Yang M Z, Zou Y P, Han C P 2015 Opt. Prec. Eng. 23 3322
Google Scholar
[11] Xu L, Wang J, Liu J G, Gao M G, Lu Y H, Liu W Q, Wei X L, Zhang T S, Chen 徐H, Liu Z M 2007 J. Atmosph. Environ. Opt. 2 218
Google Scholar
徐亮, 王君, 刘建国, 高闽光, 陆亦怀, 刘文清, 魏秀丽, 张天舒, 陈华, 刘志明 2007 大气与环境光学学报 2 218
Google Scholar
[12] 熊伟, 施海亮, 俞能海 2015 光谱学与光谱分析 35 267
Google Scholar
Xiong W, Shi H L, Yu N H 2015 Spectrosc. Spect. Anal. 35 267
Google Scholar
[13] Saarinen P, Kauppinen J 1992 Appl. Opt. 31 2353
Google Scholar
[14] Salonen K I, Salomaa I K, Kauppinen J K 1995 Appl. Opt. 34 1190
Google Scholar
[15] 冯明春, 徐亮, 刘文清, 刘建国, 高闽光, 魏秀丽 2016 物理学报 65 168
Google Scholar
Feng M C, Xu L, Liu W Q, Liu J G, Gao M G, Wei X L 2016 Acta Phys. Sin. 65 168
Google Scholar
[16] 冯明春, 刘文清, 徐亮, 高闽光, 魏秀丽, 童晶晶, 李相贤, 陈军 2015 光学学报 35 0423002
Google Scholar
Feng M C, Liu W Q, Xu L, Gao M G, Wei X L, Tong J J, Li X X, Chen J 2015 Act. Opt. Sini. 35 0423002
Google Scholar
[17] Grohs P, Kereta Z, Wiesmann U 2016 Int. J. Wavelets Multiresolut. Inf. Process. 14 1650032
Google Scholar
[18] Dicke R H 1953 Phys. Rev. 89 472
Google Scholar
[19] Penner S S, Kavanagh R W 1953 J. Opt. Soc. Am. 43 385
Google Scholar
[20] He J, Zhang Q G 2013 Optik 124 5245
Google Scholar
[21] 徐亮, 刘建国, 高闽光, 陆亦怀, 刘文清, 魏秀丽, 张天舒, 陈华, 刘志明 2008 光谱学与光谱分析 28 1052
Google Scholar
Xu L, Liu J G, Gao M G, Lu Y H, Liu W Q, Wei X L, Zhang T S, Chen H, Liu Z M 2008 Spectrosc. Spect. Anal. 28 1052
Google Scholar

 下载:
下载:



计量
- 文章访问数: 8244
- PDF下载量: 90
- 被引次数: 0
